Lower Extremity
The Lower Limb – the Erect Gait
The bipedal upright gait not only influenced the intellectual and socio-cultural development of humans but also resulted in significant changes of the human anatomy.
In humans, the lower extremities – as seen in hominids – are locomotion and support organs, however, with a more stable and wider pelvic girdle and longer legs: The extensive pelvic bones bear the weight of the upper body and support the viscera of the abdominal cavity, enabling prolonged standing without much effort. The ability to take larger steps results in accelerated locomotion. Speed and greater range of action caused already the quadrupedal mammals to develop limbs which migrated ventrally underneath the body. The front (upper) extremities were rotated dorsally while the rear (lower) extremities were rotated ventrally. Hence, in humans the extensors of the thigh and lower leg are positioned anteriorly, whereas the upper arm and forearm extensors are located posteriorly.
The joints of the free extremity such as hip, knee, and ankle joints are supported by stable ligaments. They ensure steadiness while standing and relieve the muscle groups on buttocks, knee, and calf, which are responsible for body posture.
The stance stabilizing foot of humans – in contrast to the grasping hand-like foot in hominids – has led to less mobility of the joints, especially of the interphalangeal joints of the toes; the muscles of the foot contribute to stabilization of the foot and bracing of the plantar arch rather than enabling the fine-tuned movement of individual toes.
Pelvic Girdle
In contrast to the shoulder girdle, the pelvic girdle (Cingulum membri inferioris or pelvicum) is an almost rigid bony ring. Dorsally it consists of the sacrum (Os sacrum), which is a constituent of the spine. The sacrum is unpaired and it connects bilaterally through minimally flexible joints (Articulationes sacroiliacae) with the paired pelvic bones (Ossa coxae). The Ossa coxae form two bony half shells which join ventrally beneath the Mons pubis at the fibrocartilaginous Symphysis pubica. This resembles a bony floorless basin, where the muscles and ligaments form the pelvic floor. Each pelvic bone consists of three single bones which are connected by synostoses once growth is completed: the ilium (Os ilium, cranial), the ischium (Os ischii, caudodorsal) and the pubis (Os pubis, caudoventral).
When investigating the soft tissues of the pelvic girdle, the following picture emerges: At the ventral aspect, the inguinal region (Regio inguinalis) is positioned on either side of the Mons pubis. In the tender inguinal canal, blood vessels, muscles, and nerves (and the spermatic cord in males) descend from the interior of the abdomen to the leg (and scrotum, respectively). The pulse of the femoral artery (A. femoralis) is palpable slightly lateral to both sides of the Mons pubis in the Regio inguinalis. The actual hip region (Regio coxae) is located more laterally. Dorsally, the bilaterally curved buttocks of the gluteal region (Regio glutealis, “ho glutos”: the buttocks) rest on the bony pelvic girdle. Their convexity resulted from the adaptive evolution of the gluteal muscles due to the transition to the bipedal gait. Both buttocks are separated by a deep natal cleft (Crena ani) and the gluteal fold (Sulcus glutealis) separates them from the thigh.
Lower limb
The Pars libera membri inferioris consists of the thigh (Femur), the knee (Genu), the leg (Crus), and the foot (Pes).
The thigh (Femur) is supported by the identically named bone, which is the largest long bone of the body. In the hip joint (Articulatio coxae), the ball-like head of the Femur articulates with the hemispherical socket of the Os coxae. The range of movement of the hip joint, especially the extension, is restricted by powerful, almost centimetre-thick ligaments which are incorporated into the capsule. Since the Femur is well surrounded by muscles, one can only palpate the two (epi-)condyles (bilaterally superior to the knee) and the greater trochanter (Trochanter major) in the hip region.
In the knee region (Regio genus), the thigh bone and Tibia form the knee joint (Articulatio genus). The kneecap (Patella) is the ventral part of the knee joint and articulates with the Femur through its posterior surface. The knee is primarily a hinge joint between Femur and Tibia. In a flexed position, it also allows for a certain rotation of the leg. The posterior region of the knee, the popliteal fossa (Fossa poplitea), is soft and placable when the knee is flexed. Deep in the fossa, branches of the N. ischiadicus and the A. poplitea descend from the thigh to the leg. Therefore, the pulse of the A. poplitea is hardly palpable in a flexed position of the knee.
The lower leg (Crus, leg) is supported by a medially and anteriorly located Tibia and a laterally positioned Fibula. The head of the Fibula is easily palpable distal to the knee joint (of which the Fibula is not a part). The N. fibularis communis descends subcutaneously and dorsal to the head of the Fibula. Damage to the N. fibularis communis can occur at this point, e.g. due to pressure of a poorly padded cast.
At the transition to the foot (Pes), one can easily palpate the bilateral ankle bulges (Malleolus lateralis and medialis). The Malleolus lateralis (of the Fibula) is always positioned lower than the Malleolus medialis (of the Tibia). Just inferior and posterior to the Malleolus medialis a bundle of blood vessels, nerves and tendons descends from the dorsal aspect of the Crus to the sole of the foot. The pulse of the A. tibialis posterior is palpable near the Malleolus medialis. Both malleoli of the Tibia and Fibula articulate with the Talus, forming the ankle joint (Articulatio talocruralis). It facilitates elevation and depression of the foot. The digital extensor tendons project on the dorsum of the foot. Between them, the pulse of the A. dorsalis pedis is palpable. The skeleton of the foot includes the Tarsus, the Metatarsus, and the phalanges (Digiti). There are seven tarsal bones (Ossa tarsi), the Talus being positioned on top. Just below the Talus lies the heel bone (Calcaneus) to which the ACHILLES tendon (Tendo calcaneus) attaches at its posterior surface. At the medial side, the navicular bone (Os naviculare) lies inferior and anterior to the Talus. The above mentioned three bones form the talocalcaneonavicular joint (Articulatio talocalcaneonavicularis). It permits rotating the foot inwards (supination) and outwards (pronation). The remaining tarsal bones, the three cuneiform bones (Ossa cuneiformia) and the cuboid (Os cuboideum), are interconnected by tight and almost immobile joints. The Metatarsus is supported by five long bones, the Ossa metatarsi. Together with the tarsal bones, they form the arch of the foot. The flexible arch of the foot is mainly supported by muscles and tendons located in the sole of the foot (Planta pedis). The toes I to V (Digiti) are formed by shorter long bones, the phalanges. One starts counting at the great toe (Hallux, Digitus primus); in analogy to the thumb, the Hallux has only two phalanges.
→ Dissection Link
The musculoskeletal system is dissected in layers (stratigraphically) from superficial to deep structures.
Ventral dissection: First, the epifascial structures in the subcutaneous adipose tissue are exposed. This involves several cutaneous nerves of the Plexus lumbalis and at the distal leg around the N. fibularis superficialis from the Plexus sacralis. Then follows the dissection of the V. saphena magna ascending from the anterior aspect of the medial malleolus via the medial aspect of the knee up to the Confluens venosus subinguinalis in the groin. The fascia is opened to expose the individual muscles. Immediately beneath the inguinal ligament (Lig. inguinale), the Lacunae musculorum and vasorum together with exiting neurovascular structures are dissected. From here, the A. and V. femoralis as well as the N. saphenus are traced to their entrance into the adductor canal (Canalis adductorius). Next, the origin and the branches of the A. profunda femoris, the main blood vessel supplying the thigh, are dissected. Finally, the individual joints (e.g. knee joint) are exposed.
Dorsal dissection: After exposure of the epifascial cutaneous nerves from the Plexus sacralis, the V. saphena parva is traced from the posterior aspect of the lateral malleolus to its confluence in the popliteal fossa. Next, the opening of the fascia displays the individual muscles. In the gluteal region, the M. gluteus maximus is exposed and reflected, followed by the display of the deep muscles of the gluteal region. The Regio glutealis with pathways is dissected. The N. ischiadicus is traced to its divergence and from there the N. tibialis and the N. fibularis communis with its branches are traced to the foot. The popliteal fossa is dissected including blood vessels. In the leg, the neurovascular pathways are traced along the A. tibialis anterior and posterior to the foot. After the removal of the plantar aponeurosis on the Planta pedis, the individual layers of the short foot muscles are exposed and the associated pathways are visualised.
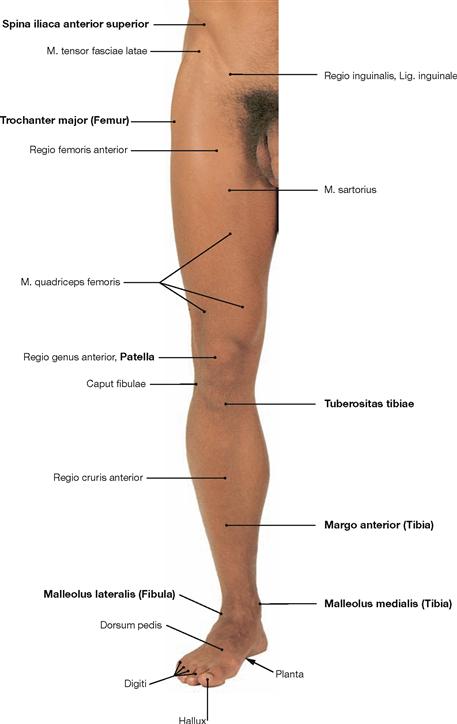
Surface anatomy
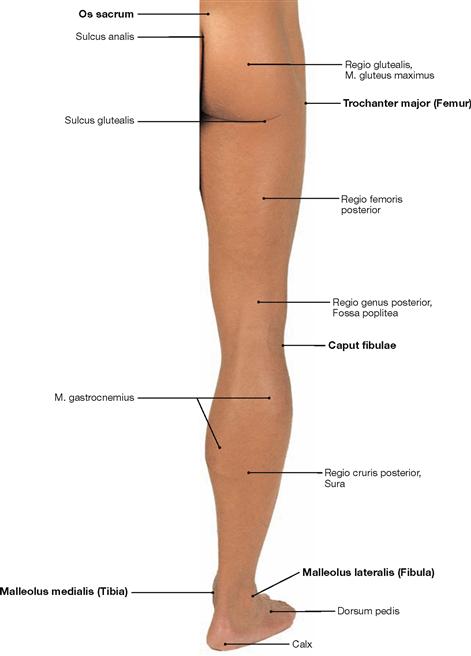
Skeleton
Skeleton of the lower extremity
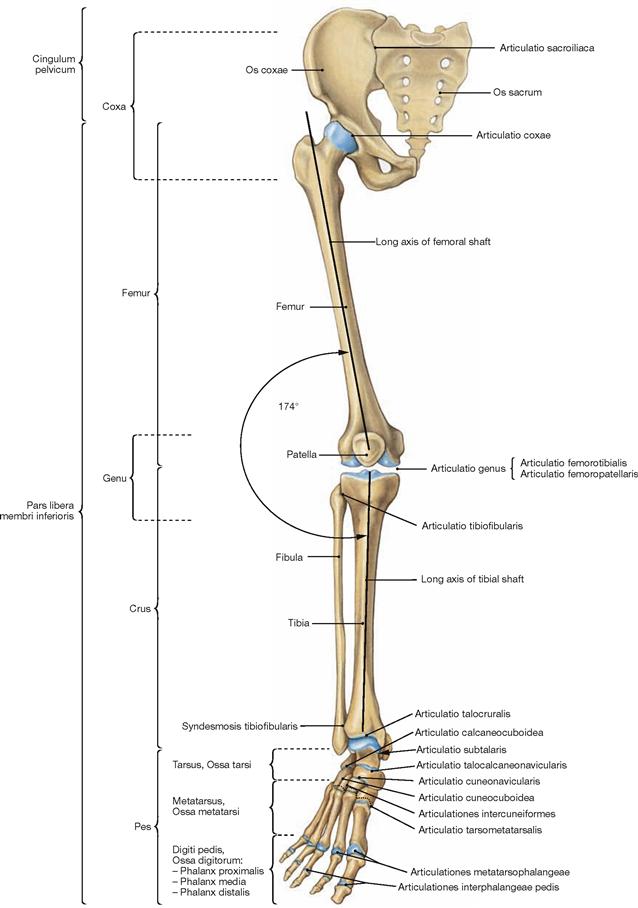
Fig. 4.3 Bones and joints of the lower extremity, Membrum inferius, right side; ventral view.
Whereas the shoulder girdle consists of two bones (Scapula and Clavicula), the pelvic girdle (Cingulum pelvicum) is formed by two hip bones (Os coxae) and the sacrum (Os sacrum). Thigh and leg form a laterally open angle of 174°, referred to as Q-angle.
In the knock-knee deformity (Genu valgum) the Q-angle is smaller, in the bowleg deformity (Genu varum) it is larger. For the development of the lower extremity → pages 132 and 133.
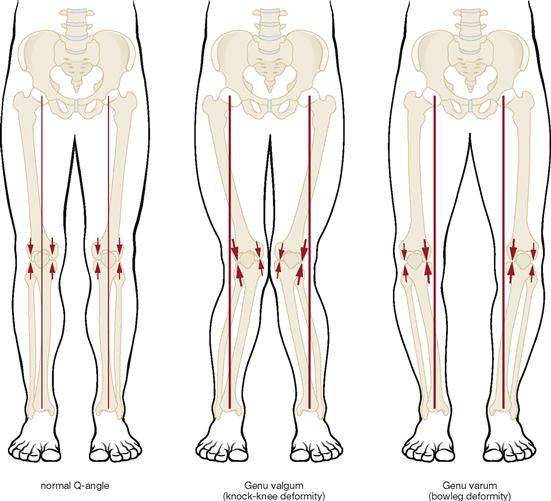
Fig. 4.4 Mechanical axis of the lower extremity (MIKULICZ’s line). (according to [1]).
Normally, the great joints of the lower extremity are positioned on a virtual straight line, the mechanical axis of the lower extremity. This axis connects the centre of the femoral head with the middle of the malleolar mortice of the ankle joint.
In the knock-knee deformity (Genu valgum), the knee is shifted medially away from the mechanical axis, in the bowleg deformity (Genu varum), it is shifted laterally.
The size of the arrows depicts the stress on the medial and lateral parts of the joint in relation to the mechanical axis.
Pelvis
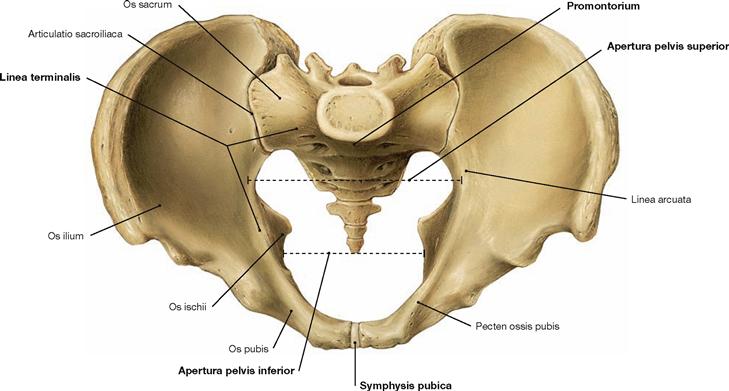
Fig. 4.5 Pelvis, Pelvis; ventral cranial view.
Sacro-iliac joint (Articulatio sacroiliaca) and pubic symphysis (Symphysis pubica) connect the two hip bones (Ossa coxae) and the sacrum (Os sacrum). The resulting stable ring formation encompasses the viscera with its iliac bones and transfers the weight of the body to the lower extremities.
The Linea terminalis begins at the pubic symphysis with the Pecten ossis pubis and continues through the Linea arcuata to the promontory (Promontorium). The Linea terminalis encircles the pelvic inlet (Apertura pelvis superior) and separates the cranial false (large) pelvis (Pelvis major) from the caudal true (small) pelvis (Pelvis minor). The promontory is the part of the vertebral column that protrudes farthest into the pelvic inlet. The pelvic outlet (Aperura pelvis inferior) is confined by the inferior margin of the pubic symphysis anteriorly, the ischial tuberosities laterally, and the tip of the coccyx posteriorly.

Fig. 4.6 and Fig. 4.7 Pelvis, Pelvis, of a woman (→ Fig. 4.6) and of a man (→ Fig. 4.7).
The shape of the pelvis shows differences between the sexes. In men, the pelvic inlet is rather heart-shaped. The smaller pubic angle is referred to as Angulus subpubicus (→ Fig. 4.41). In women, the pelvic inlet is transverse oval in shape. In addition, the inferior pubic angle (Arcus pubis, → Fig. 4.42), the distance between the ischial tuberosities, and the wings of ilium are larger than in men.
The following inner diameters are used to determine the width of the pelvic inlet: the obstetric conjugate diameter (Diameter vera) between the posterior aspect of the pubic symphysis and the promontory, the transverse diameter (Diameter transversa) between the most lateral points of the Linea terminalis on both sides, and the oblique diameter (Diameter obliqua I and II) which connects the Articulatio sacroiliaca of each side with the corresponding most distal point on the Linea terminalis.
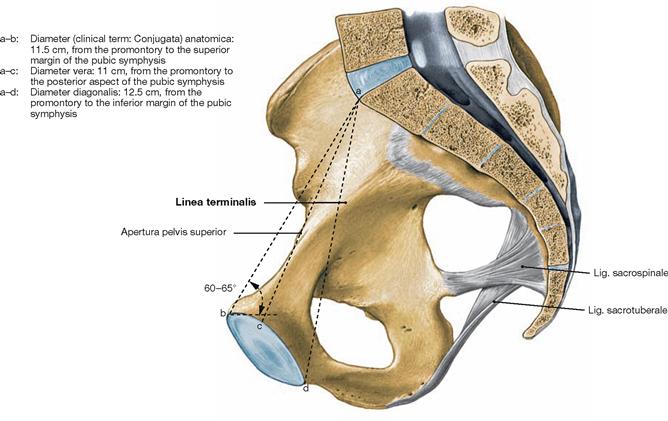
Fig. 4.8 Pelvis, Pelvis, of a woman; medial view; median section with illustration of the diverse straight inner diameters and their normal length which may, however, show interindividual variations.
The most important is the Diameter vera connecting the posterior aspect of the pubic symphysis and the promontory.
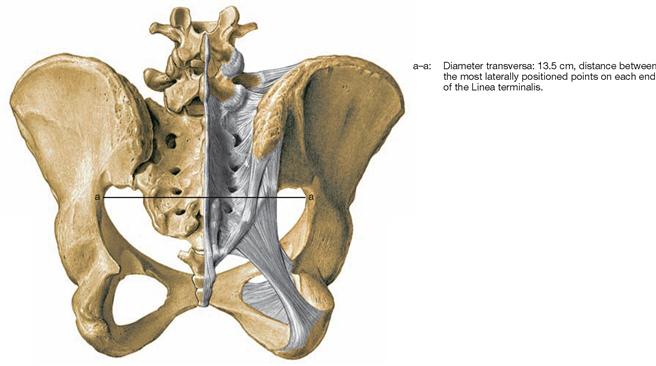
Fig. 4.9 Pelvis, Pelvis, of a woman with measurements; dorsal view.
Another internal diameter with a certain significance is the transverse diameter (Diameter transversa). The different external diameters (Distantiae), however, are of insignificant practical relevance and therefore not shown.
Hip bone
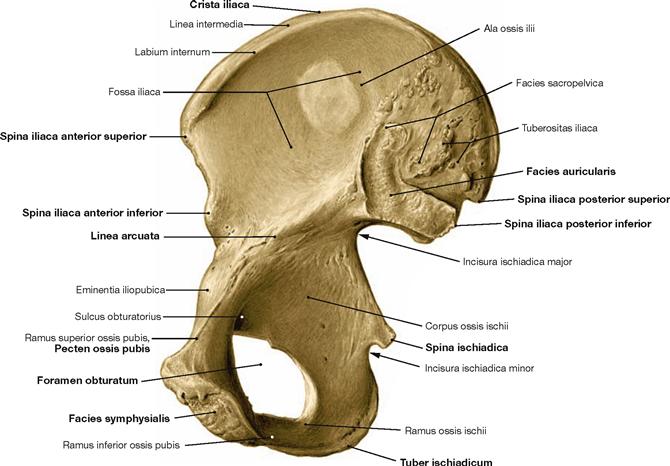
Fig. 4.10 Hip bone, Os coxae, right side; medial view.
The hip bone consists of three parts, the ilium (Os ilium), ischium (Os ischium), and pubis (Os pubis). The ilium forms the false pelvis, ischium and pubis form the bony ring around the obturator foramen from posterior and anterior, respectively. The Facies auricularis serves as articular surface for the sacro-iliac joint. The Discus interpubicus is attached to the Facies symphysialis.
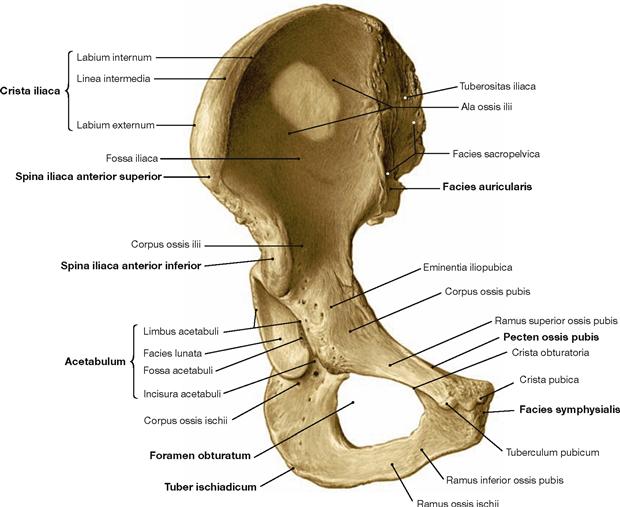
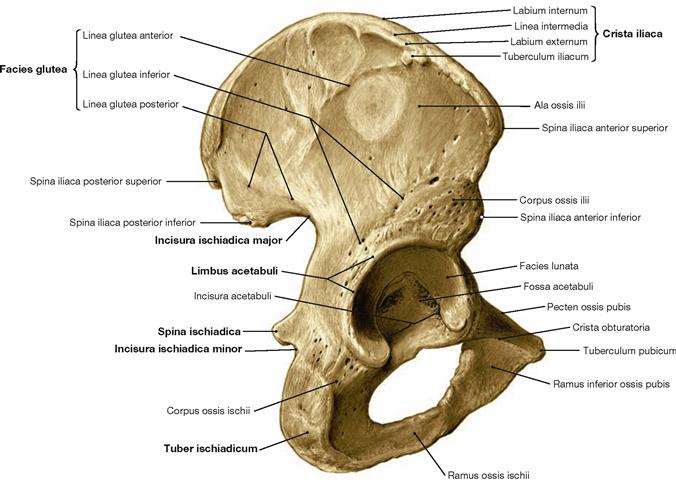
Fig. 4.12 Hip bone, Os coxae, right side; dorsolateral view.
All three parts of the hip bone, namely the ilium (Os ilium), the ischium (Os ischium) and the pubis (Os pubis), contribute to the formation of the acetabular fossa (Acetabulum).
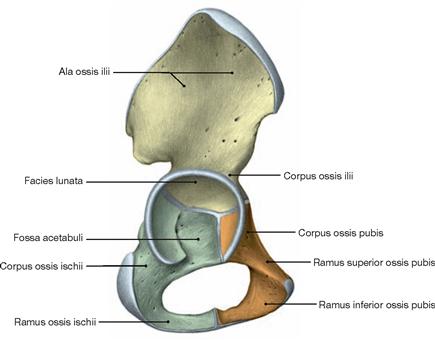
Fig. 4.13 Hip bone, Os coxae, of a 6-year-old child, right side; lateral view.
The three parts of the hip bone (Os ilium, Os ischium, Os pubis) are linked by a Y-shaped cartilaginous synchondrosis in the Acetabulum. This cartilaginous synchondrosis ossifies between the age of 13 to 18.
Thigh bone
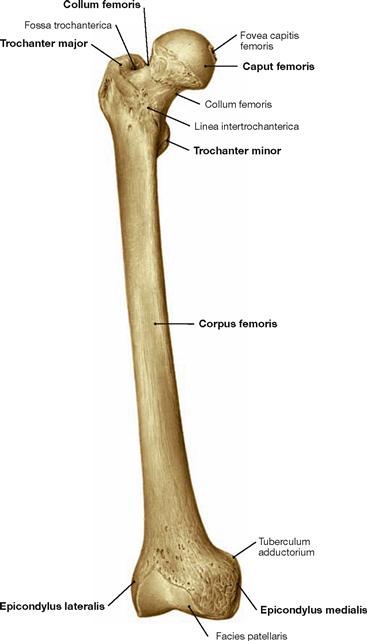
Fig. 4.14 Thigh bone, Femur, right side; ventral view.
Proximal at the femoral shaft the Trochanter major is positioned laterally and the Trochanter minor dorsomedially.
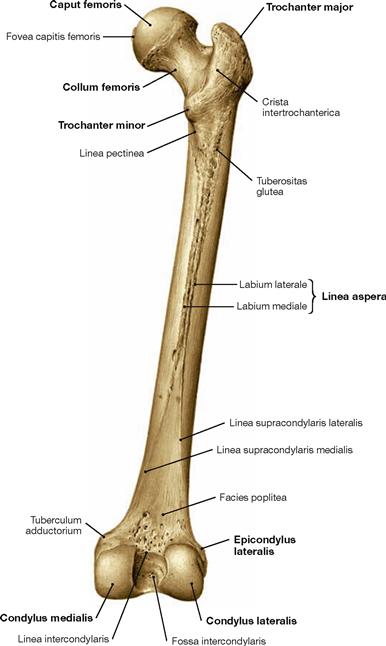
Fig. 4.15 Thigh bone, Femur, right side; dorsal view.
The Linea aspera serves as apophysis for the origin of the M. quadriceps femoris as well as for the insertion of several muscles of the adductor group.
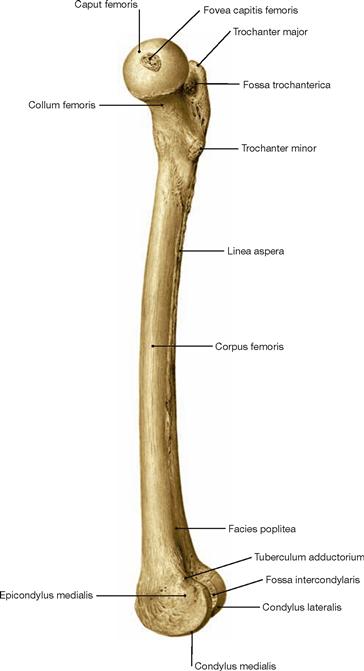
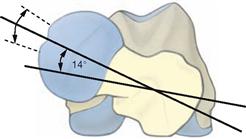
Fig. 4.17 Thigh bone, Femur, right side; proximal view; the proximal and distal ends of the femur are projected on top of each other.
The femoral neck is rotated anteriorly by 12–14° against the axis connecting both femoral condyles (transverse axis of the femoral condyles). This is referred to as torsion angle of the femur. In infants, this angle is approximately 30º.
If the torsion angle of the femur is larger, the leg is medially rotated and the toes point inwards during walking. If the torsion angle of the femur is smaller than 12°, the toes point outwards.
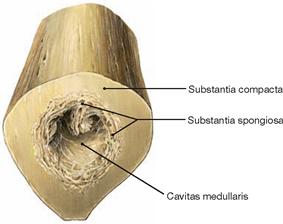
Fig. 4.18 Thigh bone, Femur, right side; cross-section of the femoral shaft at mid-level; distal view.
The outer layer of solid Substantia compacta is followed by an inner layer of Substantia spongiosa and the central medullary cavity (Cavitas medullaris) which contains the bone marrow.
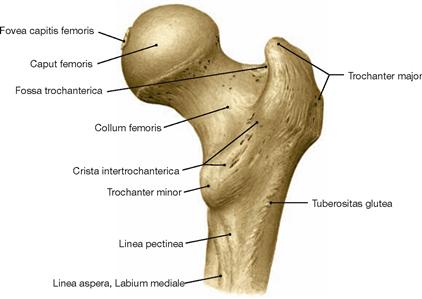
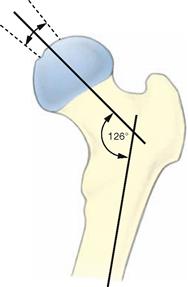
Fig. 4.20 Proximal end of the femur, Femur, right side, with illustration of the angle of inclination of the femur (neck-shaft angle)
The femoral neck forms an angle of 126° with the femoral shaft. This angle is referred to as the caput-collum-diaphyseal angle or CCD angle. In the newborn, the CCD angle measures 150°. An increased CCD angle results in a Coxa valga, a decreased CCD angle causes a Coxa vara.
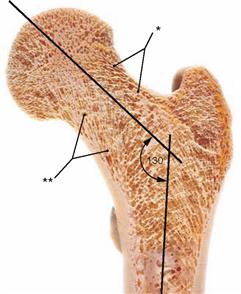
Fig. 4.21 Proximal end of the femur, Femur, right side, with illustration of the spongiosa structure in the case of an increased angle of inclination (neck-shaft angle) (Coxa valga). Section at the level of the torsion angle of the femur.
The spongiosa trabeculae are trajectorial, i.e. they align with the lines of maximal traction and compression forces (the so-called trajectories). Coxa valga causes higher compression forces and leads to a reinforcement of the medial spongiosa trabeculae (**) and at the same time to a reduction of the lateral spongiosa trabeculae (*).
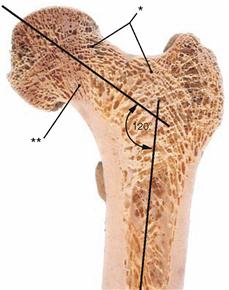
Fig. 4.22 Proximal end of the femur, Femur, right side, with illustration of the spongiosa structure in the case of a decreased angle of inclination (neck-shaft angle) (Coxa vara). Section at the level of the torsion angle of the femur.
In Coxa vara, increased traction forces cause a reinforcement of the lateral spongiosa trabeculae (*) and at the same time a reduction of the medial spongiosa trabeculae (**). As a result of an increased bending stress, the corticalis at the inner side of the femoral neck is thickened.
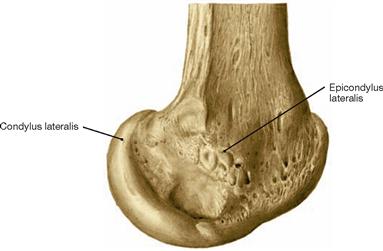
Fig. 4.23 Distal end of the femur, Femur, right side; lateral view.
To understand the flexion-extension movement in the knee joint (→ Fig. 4.69) knowledge about the articular surfaces of the femoral condyles is important. In relation to the axis of the femoral shaft the articular surfaces are positioned dorsally (retroposition). In addition, the curvature of the femoral condyles is more pronounced posteriorly (smaller radius of curvature) than anteriorly (larger radius of curvature) resulting in a spiral curvature. This phenomenon is more distinct in the medial than in the lateral condyle (→ Fig. 4.100).
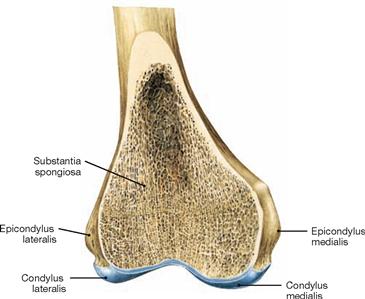
Fig. 4.24 Distal end of the femur, Femur, right side; frontal section at the level of the joint bodies; ventral view.
Tibia
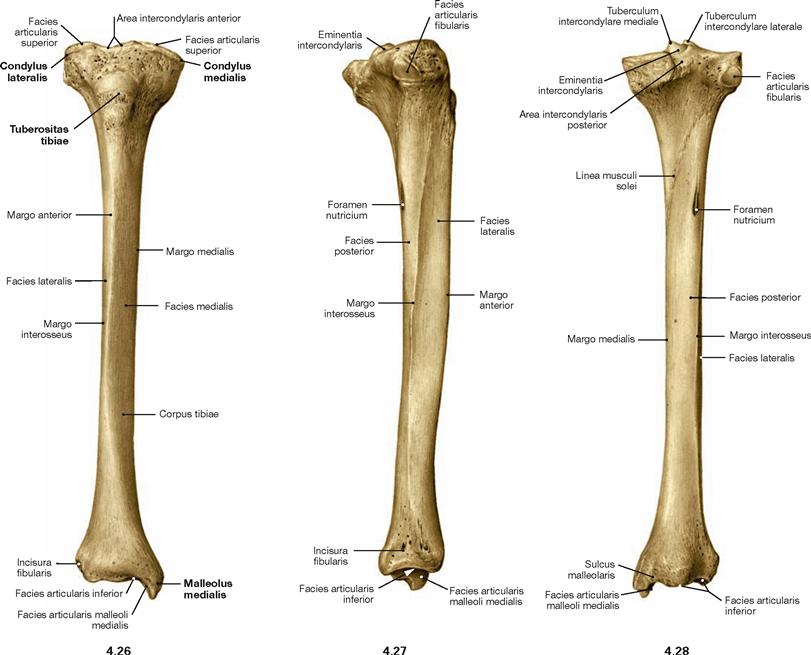
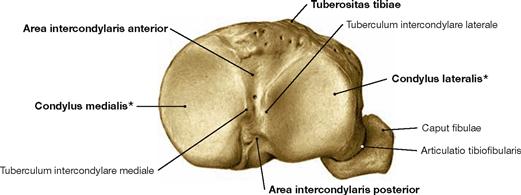
Fig. 4.26 to Fig. 4.28 Tibia, Tibia, right side; ventral (→ Fig. 4.26), lateral (→ Fig. 4.27), and dorsal (→ Fig. 4.28) views.
The proximal articular surface is shifted dorsally from the axis of the tibial shaft (retroposition). In addition, the articular surface is tilted dorsally by 3°–7° (retroversion). The retroversion is more pronounced at the medial condyle than at the lateral condyle and is here especially distinet at the medial rim of the articular surface.
Fibula
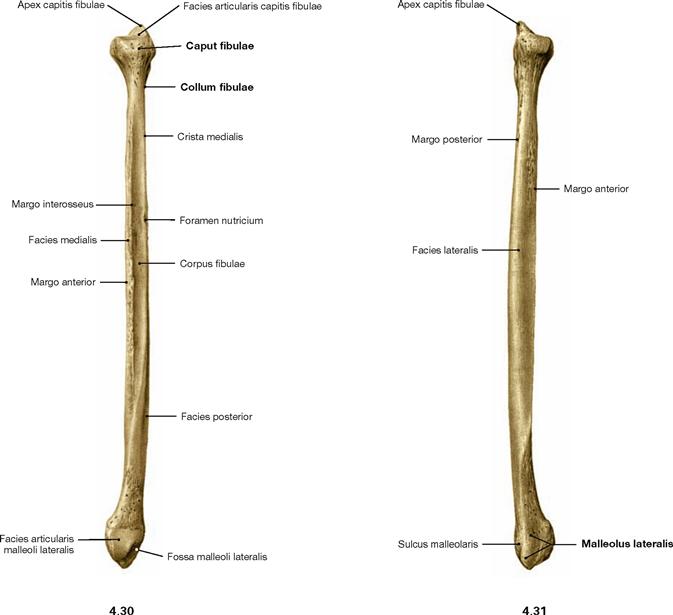
Fig. 4.30 and Fig. 4.31 Fibula, Fibula, right side; medial (→ Fig. 4.30) and lateral (→ Fig. 4.31) views.
When positioning an isolated fibula, orientation is given by the fact that the articular surfaces of the fibular head and of the malleus both point medially.
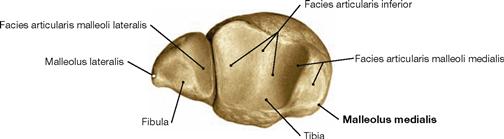
Skeleton of the foot
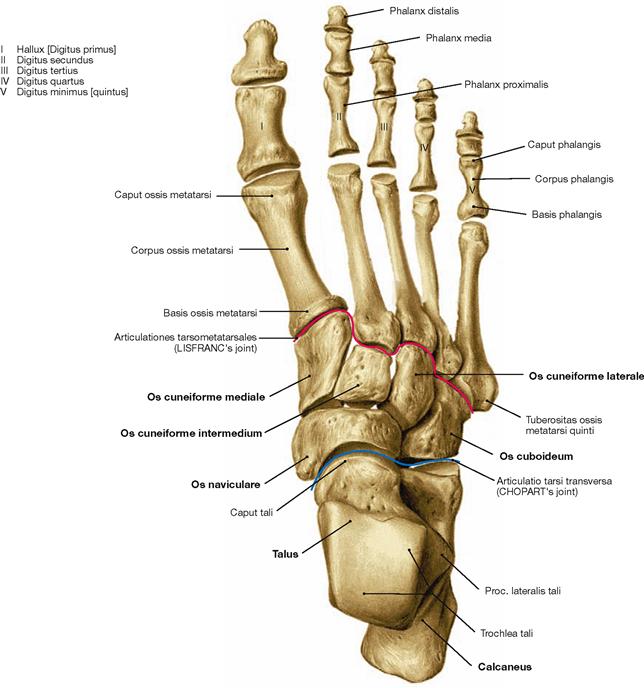
Fig. 4.33 Skeleton of the foot, Ossa pedis, right side; dorsal view.
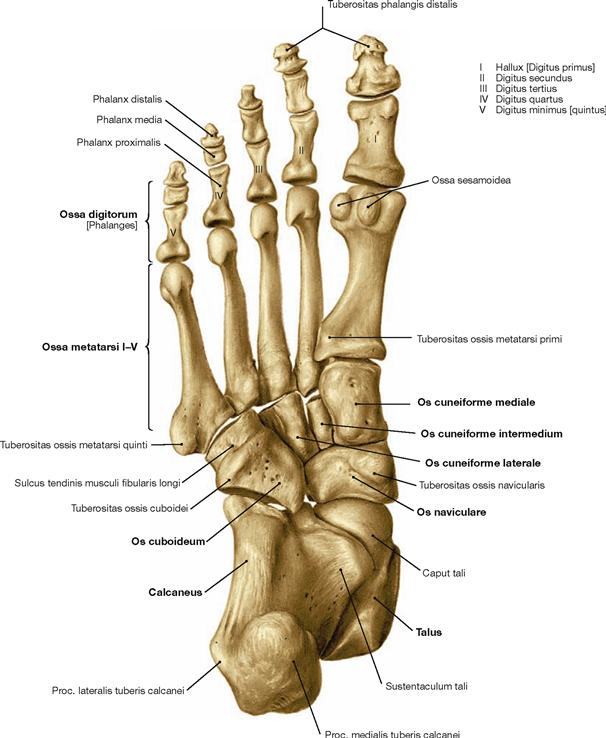
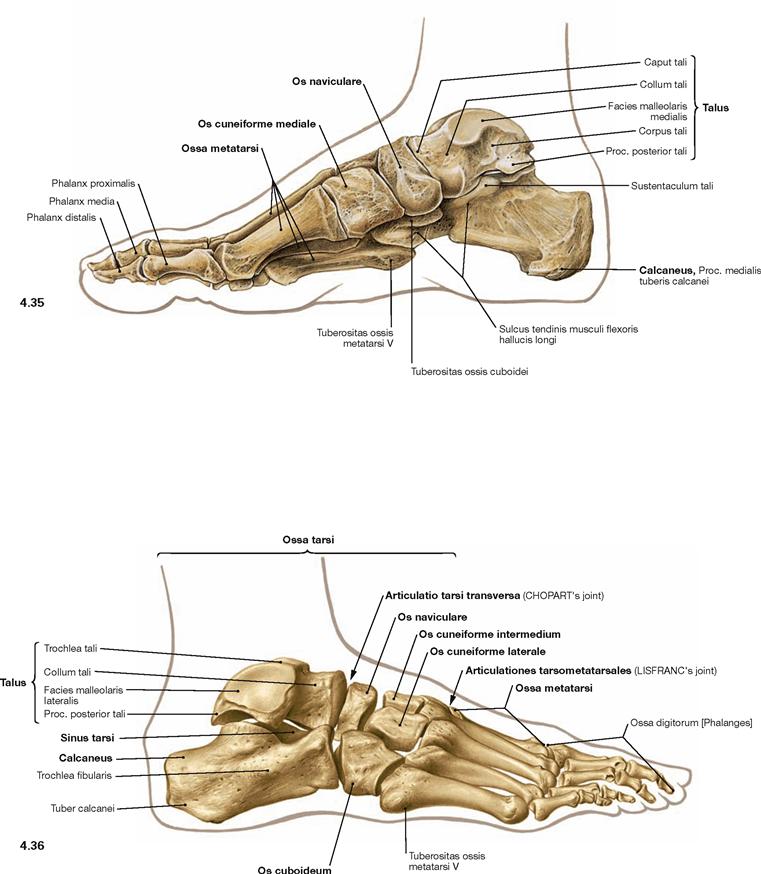
The foot (Pes) is organised in Tarsus with Ossa tarsi, Metatarsus with Ossa metatarsi, and toes (Digiti) which consist of several phalanges. The Tarsus comprises the Talus, the Calcaneus, the navicular (Os naviculare), the cuboid (Os cuboideum), and the three cuneiform bones (Ossa cuneiformia). Clinically, the forefoot is distinguished from the hindfoot. Both are separated by the articular line in the Articulationes tarsometatarsales.
Talus and calcaneus
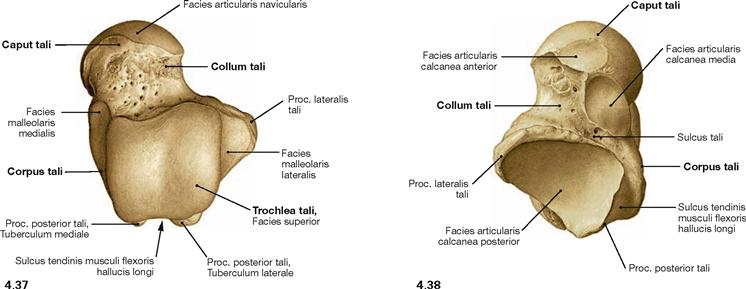
Fig. 4.37 and Fig. 4.38 Talus, Talus, right side; dorsal (→ Fig. 4.37) and plantar views (→ Fig. 4.38).
The trochlea is broader at its posterior aspect than at its anterior aspect.
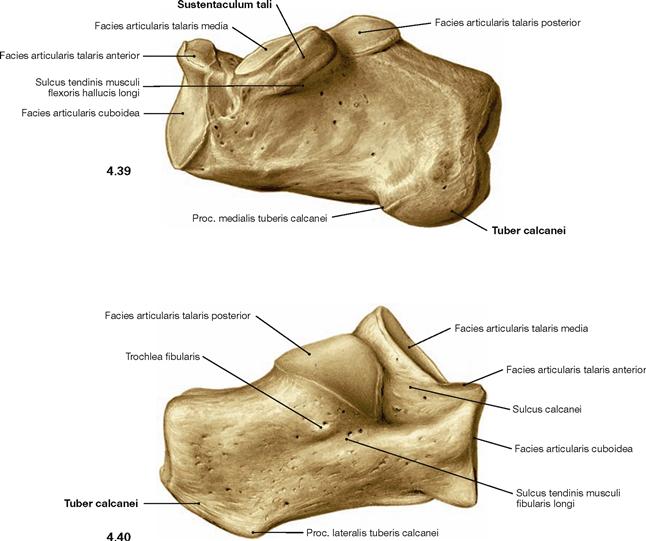
Ligaments of the pelvis
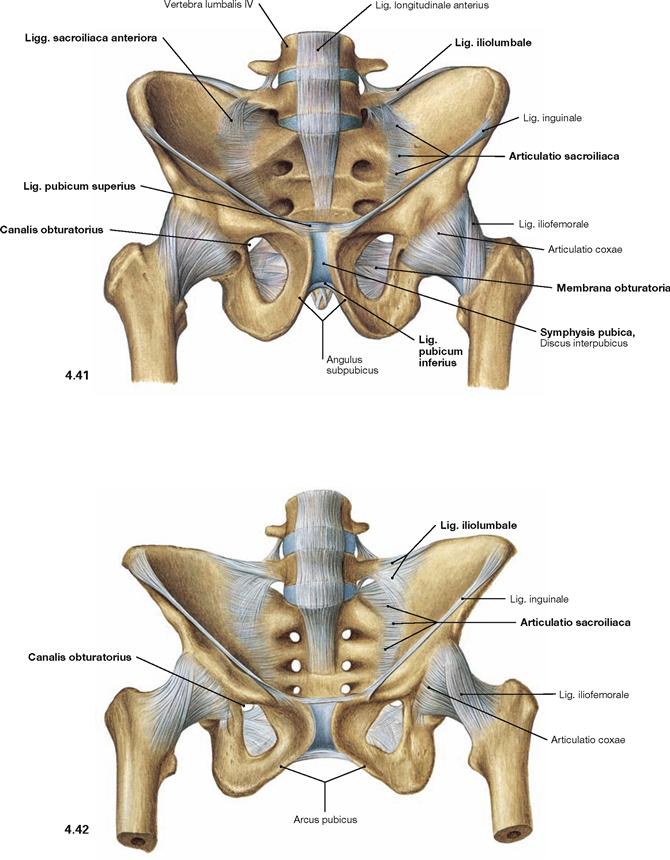
Fig. 4.41 and Fig. 4.42 Joints and ligaments of the male (→ Fig. 4.41) and the female pelvis (→ Fig. 4.42); ventral view.
The pelvic girdle (Cingulum pelvicum) is a ring-shaped bony construction created by both the dorsal amphiarthroses of the sacro-iliac joints (Articulationes sacroiliacae) and by the ventrally located pubic symphysis (Symphysis pubica). Each sacro-iliac joint is stabilised by the Ligg. sacroiliaca anteriores ventrally, and by the Lig. iliolumbale superiorly. The latter connects the Proc. costalis of the lumbar vertebrae IV and V with the Crista iliaca. These strong ligaments only allow small tilting movements of the pelvis of about 10°.
The pubic symphysis is bridged superiorly by the Lig. pubicum superius, and inferiorly by the Lig. pubicum inferius.
In both sexes the Foramen obturatum is almost completely closed by the Membrana obturatoria which only leaves the Canalis obturatorius as a passageway for the neurovascular bundles to the inner side of the thigh (A./V. obturatoria, N. obturatorius).
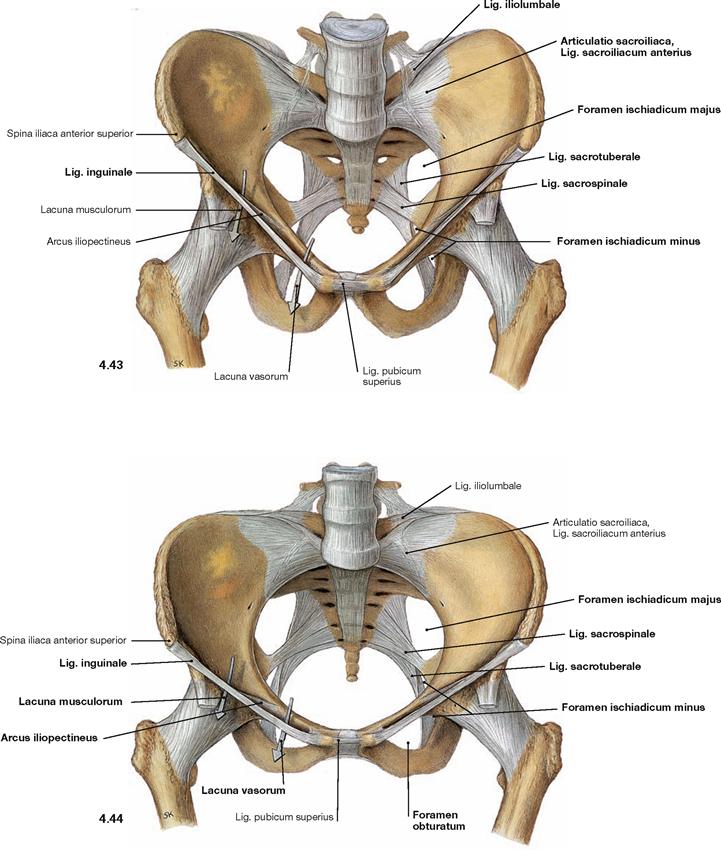
Fig. 4.43 and Fig. 4.44 Joints and ligaments of the male (→ Fig. 4.43) and the female pelvis (→ Fig. 4.44); ventral cranial view.
The almost horizontally oriented Lig. sacrospinale connects the sacrum with the Spina ischiadica, dorsal of which the Lig. sacrotuberale courses obliquely to the Tuber ischiadicum. Both ligaments complement the Incisura ischiadica major and minor to form the Foramen ischiadicum majus and minus. These openings constitute important passageways for blood vessels and nerves of the Plexus sacralis to the gluteal region (Regio glutealis). The space beneath the inguinal ligament (Lig. inguinale) is divided by the Arcus iliopectineus into the lateral Lacuna musculorum and the medial Lacuna vasorum (→ Fig. 4.177) through which the neurovascular structures course to the anterior side of the thigh.
Fig. 4.45 and Fig. 4.46 Joints and ligaments of the female pelvis; dorsal (→ Fig. 4.45) and caudal views (→ Fig. 4.46).
On the dorsal side, the sacro-iliac joint is stabilised by the Ligg. sacroiliaca posteriora and interossea. Due to the strong ligaments on the posterior side of the pelvis, only small tilting movements of up to 10° are possible.
The almost horizontally oriented Lig. sacrospinale connects the sacrum with the Spina ischiadica, dorsal of which the Lig. sacrotuberale courses obliquely to the Tuber ischiadicum. Both ligaments confine the Foramina ischiadica majus and minus as passageways for blood vessels and nerves of the Plexus sacralis to the gluteal region.
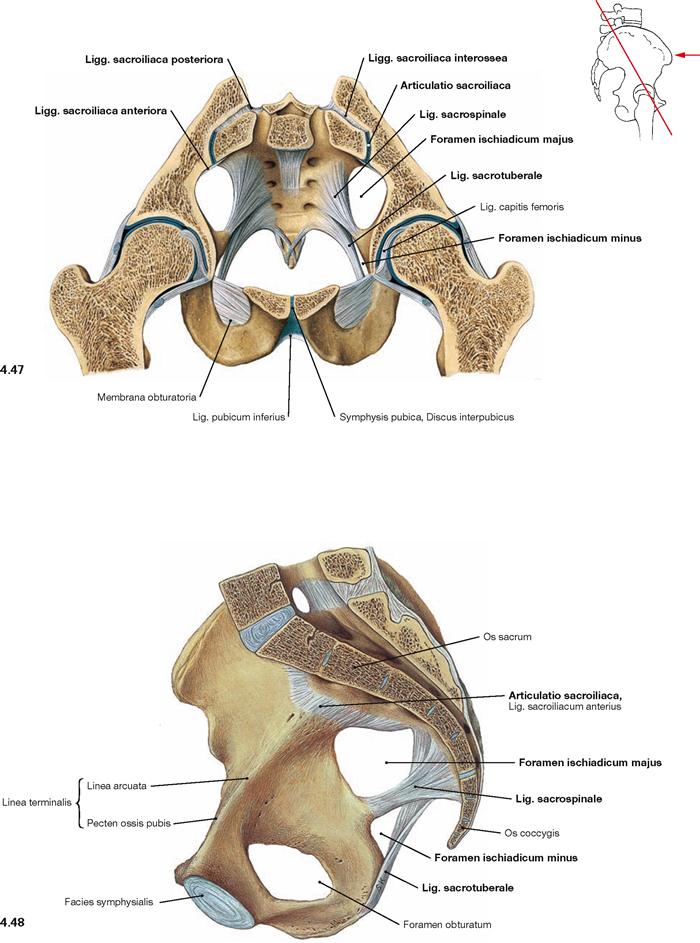
Fig. 4.47 and Fig. 4.48 Joints and ligaments of the female pelvis; oblique transverse section; ventral caudal view (→ Fig. 4.47) and median section; view from the left side (→ Fig. 4.48).
Illustrated is the sacro-iliac joint with its ligaments (Ligg. sacroiliaca anteriora, posteriora, and interossea as well as the Lig. sacrospinale, and Lig. sacrotuberale). The Lig. iliolumbale is not visible. The Lig. sacrospinale and Lig. sacrotuberale confine the Foramina ischiadica majus and minus as passageways for blood vessels and nerves of the Plexus sacralis to the gluteal region.
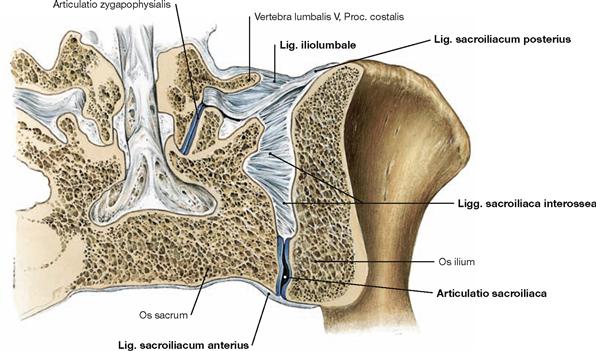
Fig. 4.49 Sacro-iliac joint, Articulatio sacroiliaca; frontal section; dorsal view.
These strong ligaments, of which the Ligg. sacroiliaca anteriora and interossea and Lig. iliolumbale are visible here, stabilise the sacro-iliac joint and enable the transmission of weight from the trunk to the pelvic girdle. In particular, the dorsal Ligg. sacroiliaca interossea and posteriora broadly connect the Sacrum and Ilium.
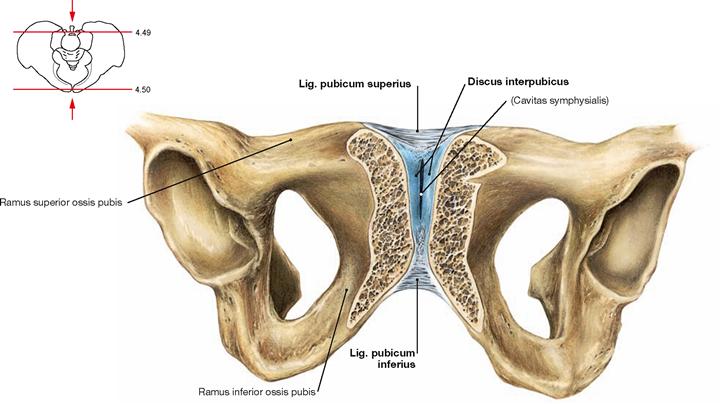
Fig. 4.50 Pubic symphysis, Symphysis pubica; oblique section; ventral caudal view.
The connection of the pubic bones is a symphysis. The Discus interpubicus consists of fibrous cartilage; only the surface area to the Facies symphysiales of both pubic bones consists of hyaline cartilage. Following the first decade of life, an oblong gap frequently forms (Cavitas symphysialis). This gap of the joint is bridged superiorly by the Lig. pubicum superius and inferiorly by the Lig. pubicum inferius.
Hip joint
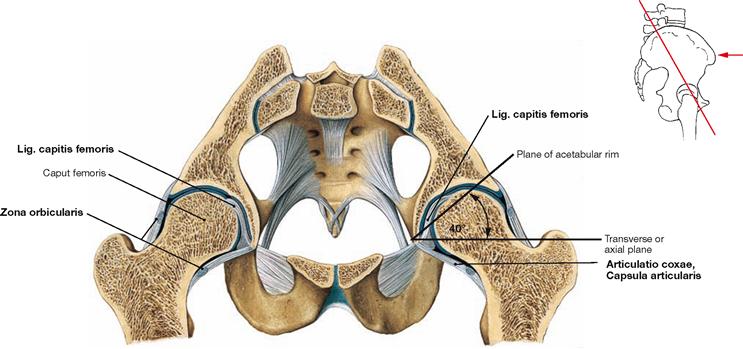
Fig. 4.51 Hip joints, Articulationes coxae; oblique transverse section; ventral cranial view.
In the hip joint, the Acetabulum of the hip bone forms the socket. Together with the Labrum acetabuli, the Acetabulum covers more than half of the femoral head (Caput femoris). Thus, the hip joint is a special form of a ball-and-socket joint referred to as cotyloid joint (Articulatio cotylica, enarthrosis). The angle between the plane of the acetabular rim and the transverse (axial) plane is 40°. The hip joint transfers the whole body weight onto the lower extremities. Therefore, the joint capsule (Capsula articularis) is reinforced by strong ligaments. Circular fibres of the joint capsule surround the femoral neck in particular on the dorsal side and are referred to as Zona orbicularis which Iigaments of the capsule also join. The Lig. capitis femoris lacks a mechanical function.
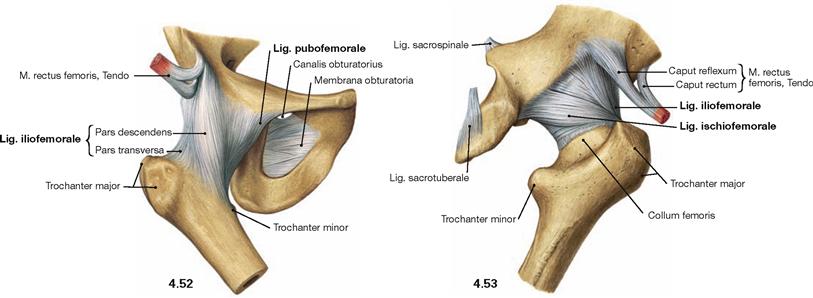
Fig. 4.52 and Fig. 4.53 Hip joint, Articulatio coxae, right side; ventral (→ Fig. 4.52) and dorsal (→ Fig. 4.53) views.
There are three major ligaments of the hip joint which surround the femoral head in a spiral manner. Their principle function is to limit the range of hip extension and to prevent the backward tilting of the pelvis:
• Lig. iliofemorale (anterior and superior): inhibits extension and adduction and, thus, supports the small gluteal muscles
• Lig. pubofemorale (anterior and inferior): inhibits extension, abduction, and lateral rotation
• Lig. ischiofemorale (posterior): inhibits extension, medial rotation, and adduction
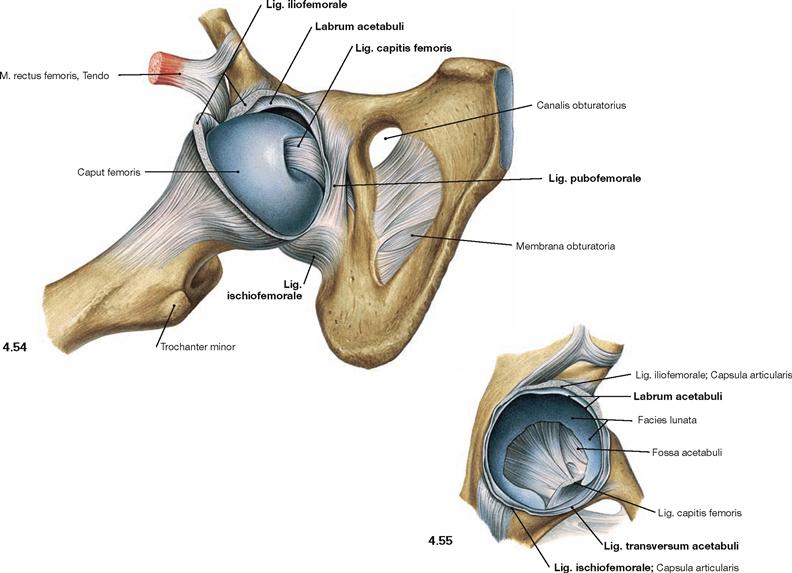
Fig. 4.54 and Fig. 4.55 Hip joint, Articulatio coxae, right side; ventral view; after opening of the capsule and partial (→ Fig. 4.54) or complete (→ Fig. 4.55) exarticulation of the femoral head.
Besides the external ligaments (Lig. iliofemorale, Lig. pubofemorale, Lig. ischiofemorale) the internally located Lig. capitis femoris is visible which lacks mechanical function in the joint. The Lig. transversum acetabuli inferiorly closes the Acetabulum, and together with the Labrum acetabuli, which also consists of fibrous connective tissue, it serves to guide the femoral head.
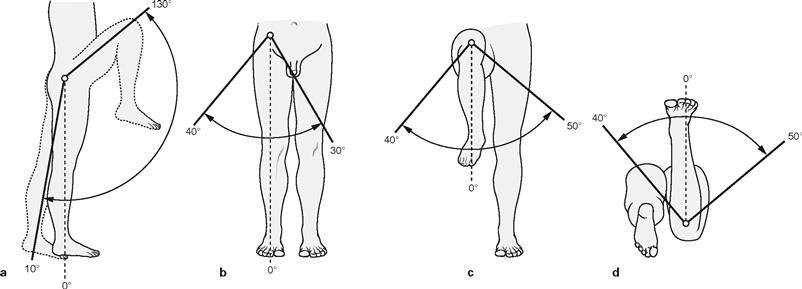
Figs. 4.56a to d Range of movement in the hip joint, Articulatio coxae. (according to [1])
The hip joint is an enarthrosis (Articulatio cotylica) which as a ball-and-socket joint possesses three axes of movement. All axes pass through the centre of the femoral head. The range of movement is limited by the strict guidance of the Acetabulum and the strong ligaments. All ligaments together restrict extension (retroversion) by enclosing the femoral head like a spiral ligamentous screw, thus enabling a stable upright position. Important for walking, the flexion (anteversion) is possible to a much higher degree and exclusively limited by soft tissues. In addition, medial and lateral rotation as well as adduction and abduction are limited by ligaments.
Range of movement:
Blood supply of the hip joint
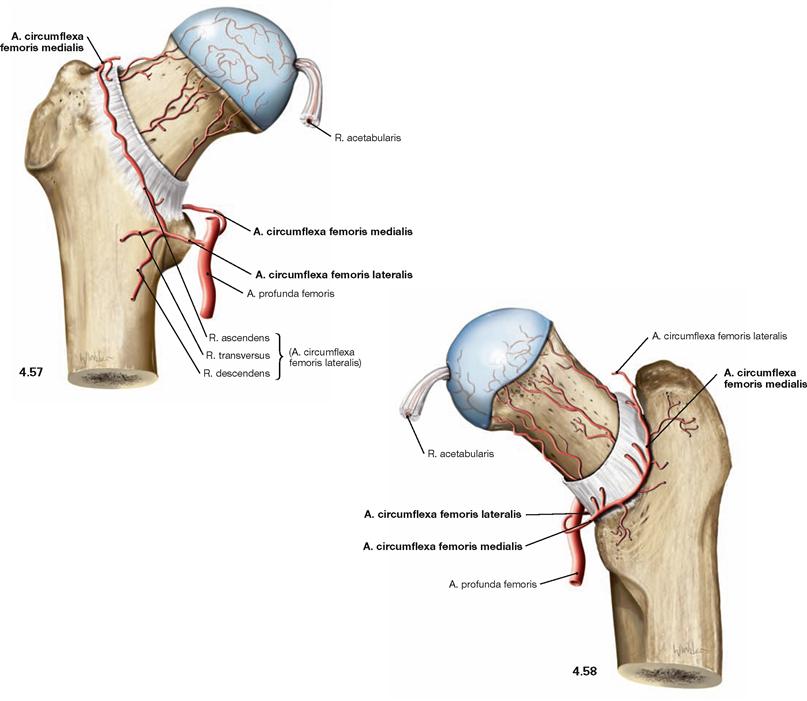
Fig. 4.57 and Fig. 4.58 Blood supply of the hip joint, right side; ventral (→ Fig. 4.57) and dorsal (→ Fig. 4.58) views.
In the adult, the A. circumflexa femoris medialis is the major blood vessel supplying the femoral head. In infants, however, the R. acetabularis (from A. obturatoria and A. circumflexa femoris medialis), which runs within the Lig. capitis femoris, provides the major part of the blood supply to the femoral head. In the adult, it supplies only one-fifth to one-third of the proximal epiphysis. However, the A. circumflexa femoris medialis supplies the femoral head and neck via several smaller branches coursing on the posterior side within the joint capsule. The A. circumflexa femoris lateralis mainly supplies the femoral neck at its anterior side. The Acetabulum is supplied from ventral and dorsal by the A. obturatoria and from cranial by the A. glutea superior.
Knee joint
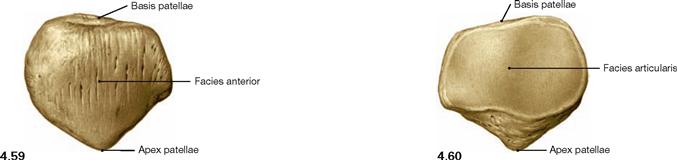
Fig. 4.59 and Fig. 4.60 Patella, Patella, right side; ventral (→ Fig. 4.59) and dorsal (→ Fig. 4.60) views.
The patella is a sesamoid bone (Os sesamoideum) within the tendon of the M. quadriceps femoris. It serves as a hypomochlion by guiding the tendon on its way to the insertion on the Tuberositas tibiae over the distal end of the femur. This results in an increase of the virtual lever arm and torque of the muscle.
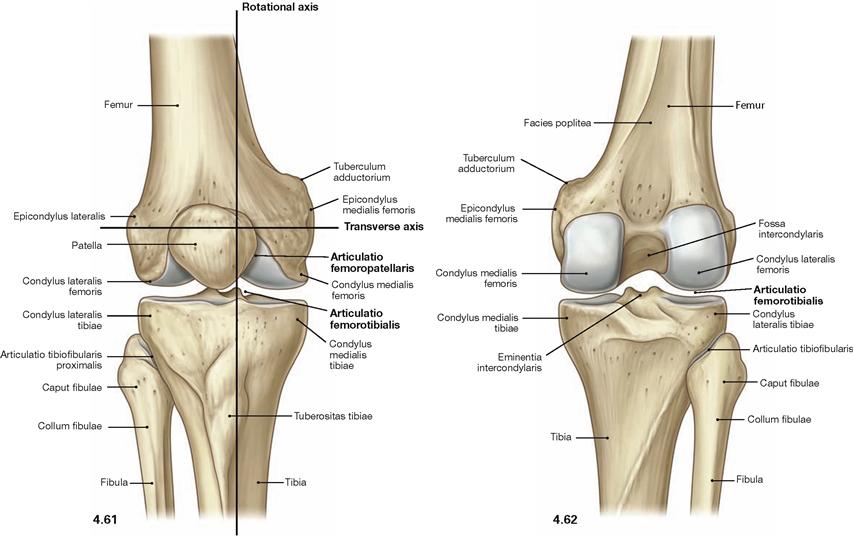
Fig. 4.61 and Fig. 4.62 Knee joint, Articulatio genus, right side; ventral (→ Fig. 4.61) and dorsal (→ Fig. 4.62) views. [10]
In the knee joint the Femur articulates with Tibia (Articulatio femorotibialis) and Patella (Articulatio femoropatellaris; → Fig. 4.209). All bones are ensheathed by a common joint capsule. In the Articulatio femorotibialis, the femoral condyles constitute the head and the upper articular surface of the Tibia (Facies articularis superior) and both tibial condyles form the socket of the joint.
The knee joint is a bicondylar joint (Articulatio bicondylaris) which functions as a pivot-hinge joint (trochoginglymus) and possesses two axes of movement. The transverse axis for extension and flexion movements extends through both femoral condyles. The longitudinal axis for rotational movements is positioned eccentrically and perpendicular through the Tuberculum intercondylare mediale. For the range of movement in the knee joint → page 276.
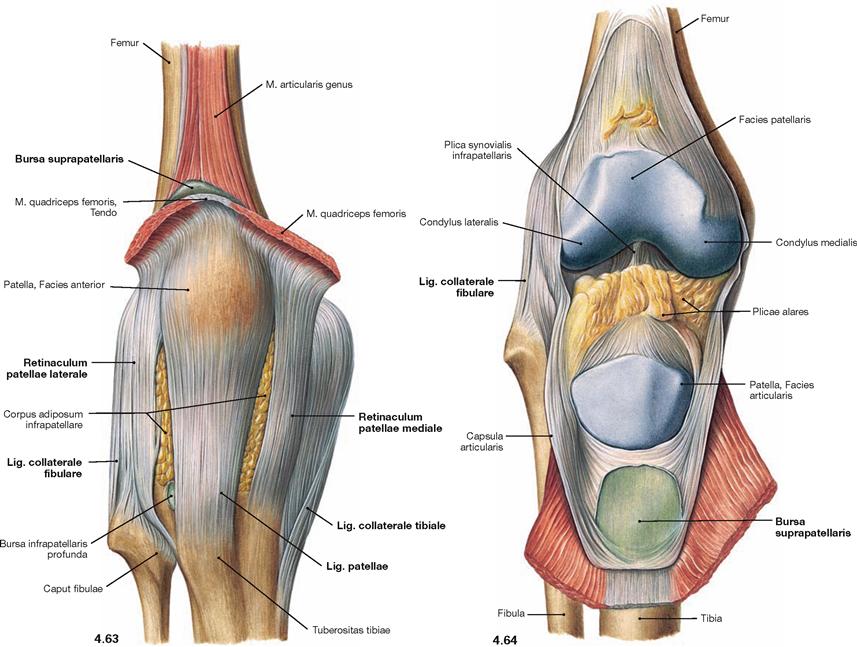
Fig. 4.63 and Fig. 4.64 Knee joint, Articulatio genus, right side; with closed joint capsule (→ Fig. 4.63), and after opening of the capsule (→ Fig. 4.64); ventral view.
The ligaments of the knee joint consist of external ligaments which support the joint from the outside, and internal ligaments which are positioned within the Capsula fibrosa. Here, the external ligaments are illustrated. They comprise the Lig. patellae as the continuation of the tendon of the M. quadriceps femoris, and the Retinacula patellae mediale and laterale. Both of these latter ligaments have superficial longitudinal and deep circular fibres and can be viewed as parts of the tendon of the M. quadriceps femoris (Mm. vasti medialis and lateralis). Medially and laterally, there are two collateral ligaments (Ligg. collateralia tibiale and fibulare) which insert in the Tibia and Fibula. The joint capsule encloses the articular surfaces. The HOFFA’s fat pad (Corpus adiposum infrapatellare) is positioned between the Capsula fibrosa and the Capsula synovialis. This adipose tissue is connected to the anterior cruciate ligament by a fold, the Plica synovialis infrapatellaris, and laterally possesses two Plicae alares. The knee joint is associated with several bursae some of which communicate with the joint capsule as shown here for the Bursa suprapatellaris.
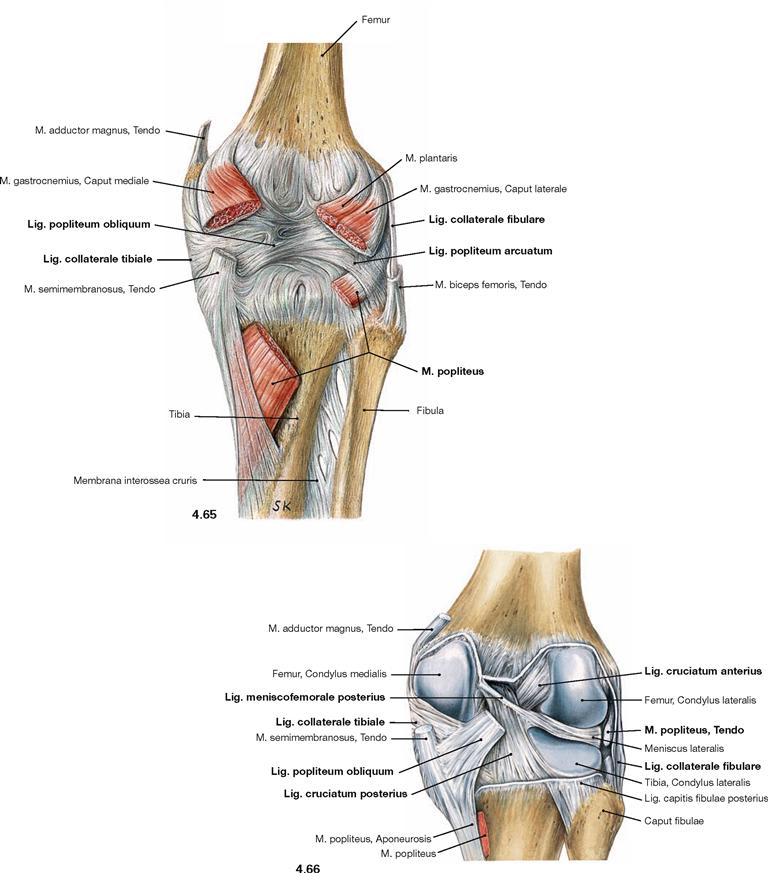
Fig. 4.65 and Fig. 4.66 Knee joint, Articulatio genus, right side; with closed joint capsule (→ Fig. 4.65), and after opening of the capsule (→ Fig. 4.66); dorsal view.
At the rear side of the knee joint, additional external ligaments support the joint capsule. The Lig. popliteum obliquum projects medially and inferiorly from the lateral femoral condyle, and the Lig. popliteum arcuatum courses in the opposite direction, thus, crossing the M. popliteus. Of the two collateral ligaments, only the Lig. collaterale tibiale is connected to the joint capsule. The Lig. collaterale fibulare is separated from the joint capsule by the tendon of the M. popliteus.
After opening of the joint capsule several internal ligaments are visible. The anterior cruciate ligament (Lig. cruciatum anterius) courses from the inner surface of the lateral femoral condyle in an anterior direction to the Area intercondylaris anterior of the Tibia. The posterior cruciate ligament (Lig. cruciatum posterius) courses in the opposite direction from the inner surface of the medial femoral condyle to the Area intercondylaris posterior of the Tibia. The Lig. meniscofemorale anterius (not visible here) and the Lig. meniscofemorale posterius connect the posterior horn of the lateral meniscus (Meniscus lateralis) anterior and posterior to the posterior cruciate ligament with the medial condyle and, thus, support the posterior cruciate ligament.
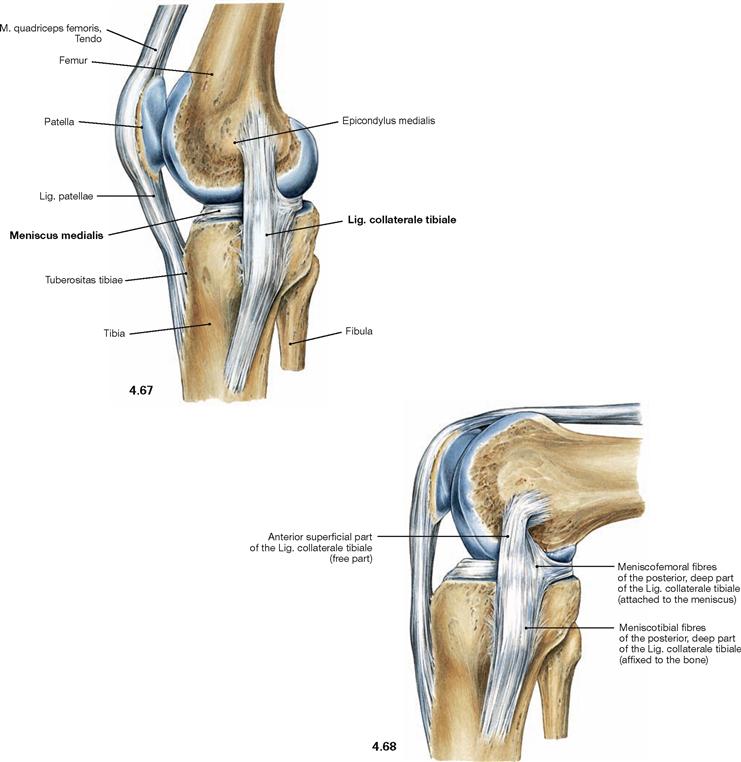
Fig. 4.67 and Fig. 4.68 Inner (medial) collateral ligament, Lig.collaterale mediale, in extension (→ Fig. 4.67), and flexion (→ Fig. 4.68); medial view.
Only the posterior fibres of the inner collateral ligament (Lig. collaterale tibiale) are connected to the Meniscus medialis. In flexion, the contortion of the ligament fixes the Meniscus medialis in its position. In contrast, the lateral collateral ligament (Lig. collaterale fibulare) is not connected to the Meniscus lateralis. Because of the larger radius of curvature of the femoral condyles in the front, the collateral ligaments are stretched in the extended knee. This position therefore does not allow for rotational movements. In a flexed position of the knee, the collateral ligaments are relaxed due to the smaller radius of curvature of the femoral condyles at the back, thus enabling rotational movements.
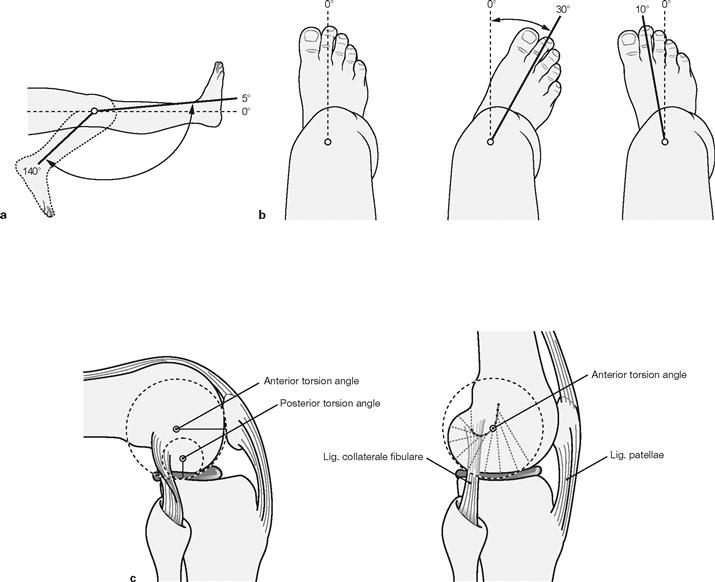
Figs. 4.69a to c Range of movement in the knee joint, Articulatio genus. (c according to [1])
The knee joint is a bicondylar joint (Articulatio bicondylaris) which functions similar to a pivot-hinge joint (trochoginglymus) and has two axes of movement. The transverse axis for extension and flexion movements runs through both femoral condyles (c). The longitudinal axis for rotational movements: projects eccentrically and perpendicular through the Tuberculum intercondylare mediale of the Tibia. Due to the smaller posterior radius of curvature of the femoral condyles the transverse axis does not remain in a constant position, but moves posteriorly and superiorly during flexion in a convex line (c). The flexion movement thus is a combined rolling and sliding movement in which the condyles roll up to 20° posteriorly and then turn in this position. Since the shape of the medial and lateral condyles of the Femur and Tibia is not identical, it is the lateral femoral condyle that predominantly rolls (similar to a rocking chair) and the medial condyle remains in its position to rotate (similar to a ball-and-socket joint). At the same time the Femur rotates slightly outwards. In the terminal phase of the extension movement, the tension of the anterior cruciate ligament also causes a forced lateral rotation of 5°–10°, during which the medial condyle even loses its contact with the medial meniscus.
The active flexion up to 120° can be increased up to 140° after pre-extension of the hamstring muscles (a). Passive flexion is possible up to 160°, limited only by soft tissues. Extension is possible up to the null-position but can be further increased passively by 5°–10°. Rotational movements are exclusively possible during flexion of the knee because the tension of the collateral ligaments during knee extension prevents rotational movements (b). Lateral rotation is possible to a larger extent than medial rotation because the cruciate ligaments twist around each other during medial rotation. Abduction and adduction are almost completely prevented by the strong collateral ligaments.
Range of movement:
Cruciate ligaments
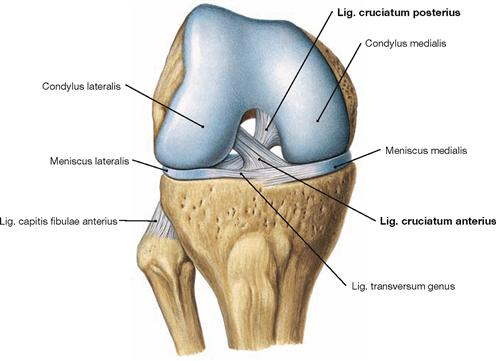
Fig. 4.70 Knee joint, Articulatio genus, right side, in 90°-flexed position; ventral view; after removal of the joint capsule and the collateral ligaments.
The most important inner ligaments are the two cruciate ligaments. The anterior cruciate ligament (Lig. cruciatum anterius) courses from the inner surface of the lateral femoral condyle in an anterior direction to the Area intercondylaris anterior of the Tibia (from a superior posterior lateral to an inferior anterior direction). The posterior cruciate ligament (Lig. cruciatum posterius) courses in an opposite direction from the inner surface of the medial femoral condyle to the Area intercondylaris posterior of the Tibia (from a superior anterior medial to a posterior inferior direction). Although the cruciate ligaments are positioned within the fibrous joint capsule (intra-articular) they are outside the Capsula synovialis and thus extrasynovial.
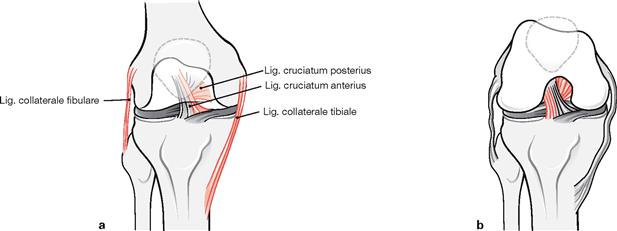
Figs. 4.71a to b Stabilisation of the knee joint, Articulatio genus, right side, through collateral and cruciate ligaments in extension (a), and flexion (b); ventral view.
The cruciate ligaments together with the collateral ligaments form a functional unit. The collateral ligaments are tense only during extension of the knee and stabilise the knee in this position against rotational as well as abduction/adduction movements. In contrast, distinct parts of the cruciate ligaments are tense during all positions of the knee joint: the medial components during extension, and the lateral components during flexion.
Menisci
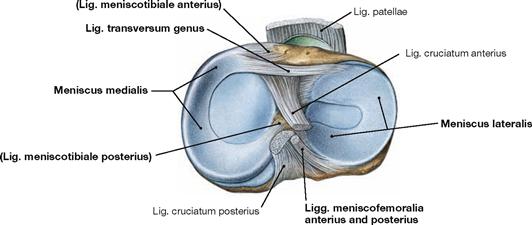
Fig. 4.72 Menisci of the knee joint, right side; cranial view.
Both menisci are roughly C-shaped and appear wedge-shaped in cross- sections. The medial meniscus is larger and anchored via the Ligg. meniscotibialia anterius and posterius to the respective Area intercondylaris of the Tibia. In addition, the medial meniscus is fixed to the medial collateral ligament. In contrast, the lateral meniscus is anchored via the Ligg. meniscofemoralia anterius and posterius to the medial femoral condyle, but it is separated from the lateral collateral ligament by the tendon of the M. popliteus (→ Fig. 4.77). The posterior horn is only indirectly and flexibly fixed to the Tibia via the M. popliteus. Anteriorly, both menisci are connected through the Lig. transversum genus. As a result, the range of movement of the lateral condyle is increased in flexion.
Both menisci are composed of fibrous carilage inside and dense connective tissue outside.
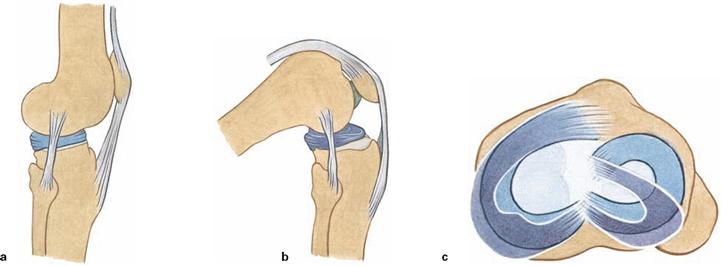
Figs. 4.73a to c Sliding range of the menisci, Menisci, during flexion.
In flexion, both menisci are pushed posteriorly over the rims of the tibial condyles. The mobility of the lateral meniscus is higher due to the reduced fixation.
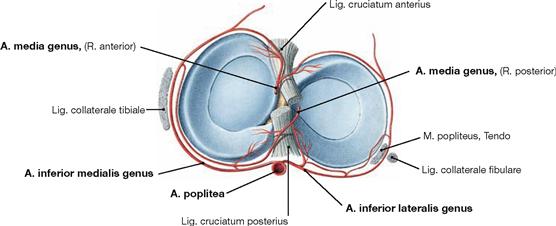
Fig. 4.74 Arterial supply of the menisci, Menisci, right side; cranial view.
The external portions of the Menisci are supplied through a perimeniscal network of blood vessels that derives from the Aa. inferiores medialis and lateralis genus and from the A. media genus (branches of the A. poplitea). The internal portions are devoid of blood vessels and are nourished by diffusion from the synovial fluid.
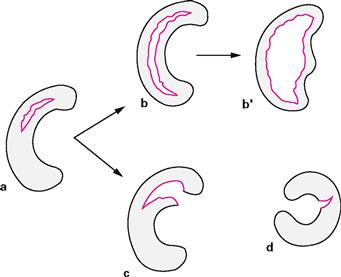
Figs. 4.75a to d Stages in the development of meniscal tears. [4]
a. development of a longitudinal tear
b. elongation of the tear from the posterior to the anterior horn and shift into the joint (“bucket handle” tear, b‘) or
c. additional radial tear (“parrot beak“; often leading to a posterior or anterior horn avulsion)
d. radial tear, lateral C-shaped meniscus most commonly affected
Knee joint
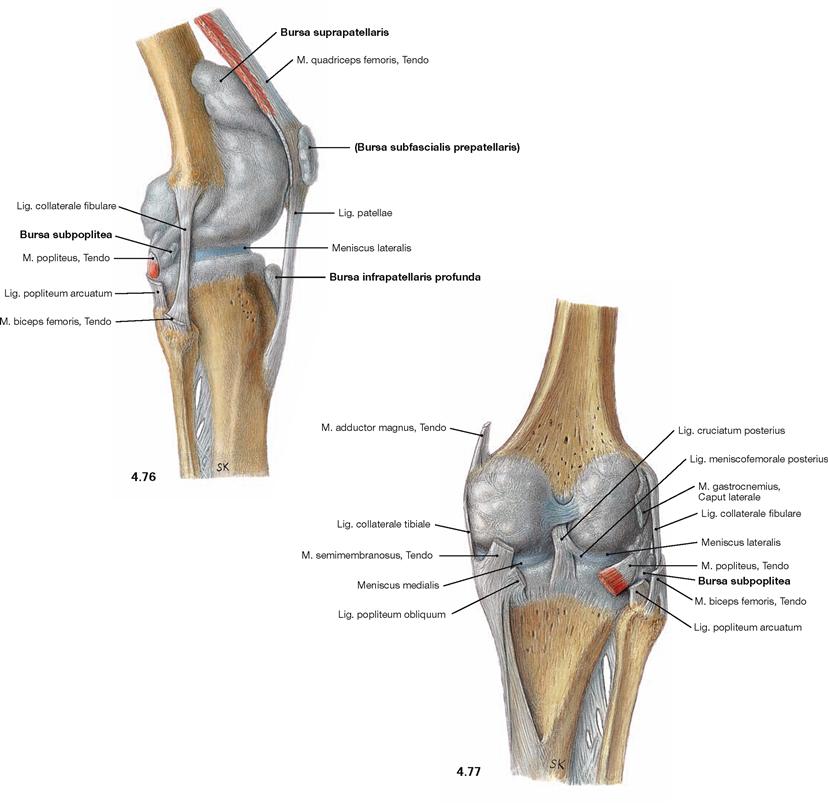
Fig. 4.76 and Fig. 4.77 Knee joint, Articulatio genus, right side, with bursae; lateral (→ Fig. 4.76) and dorsal (→ Fig. 4.77) views; illustration of the articular cavity by injection of a synthetic polymer.
The knee joint is surrounded by up to 30 bursae (Bursae synoviales). Some bursae communicate with the joint capsule, such as the Bursa suprapatellaris (anterior superior) beneath the tendon of the M. quadriceps femoris, or the Bursa subpoplitea (posterior inferior) beneath the M. popliteus. Other bursae are positioned in places with exposure to higher pressure (e.g. when kneeling) such as the Bursa prepatellaris or the Bursa infrapatellaris. Some serve as gliding surface for tendons of muscles such as the Bursa musculi semimembranosi or the Bursae subtendineae musculorum gastrocnemii medialis and lateralis (both not shown).
Knee joint, arthroscopy
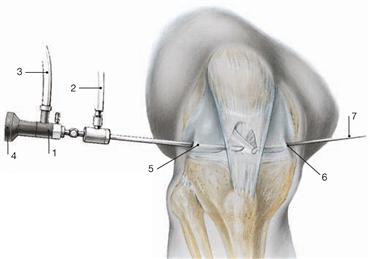
Fig. 4.78 Endoscopic examination (arthroscopy) of the knee joint.
Arthroscopy allows the minimally-invasive access to the articular cavity to assess the intrasynovial structures of the knee joint and to perform minor repairs.
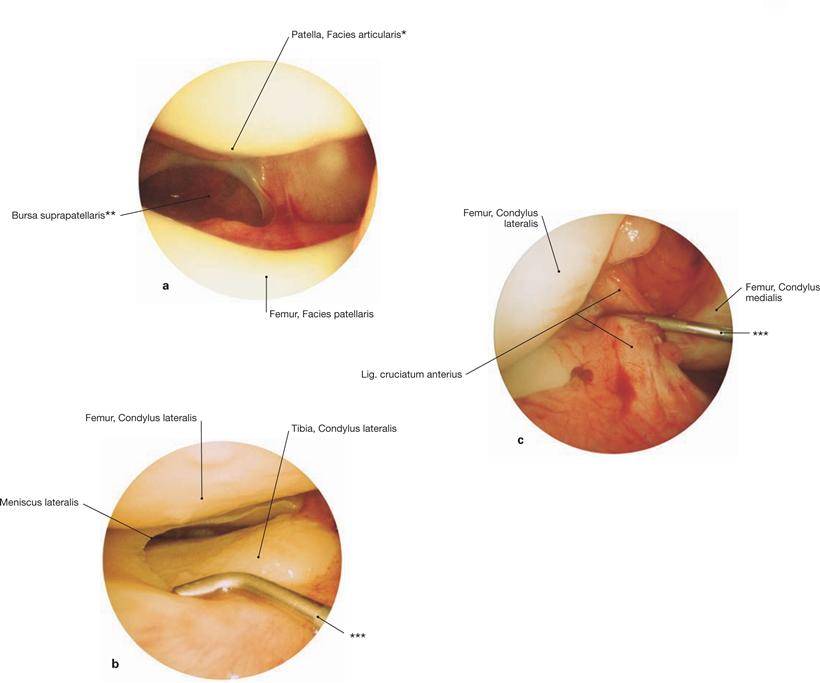
Figs. 4.79 a to c Knee joint, Articulatio genus, right side; arthroscopic images.
a. distal view in the femoropatellar joint
* patellar ridge: ridge between medial and lateral articular surfaces
** clinical term: Recessus suprapatellaris
*** groping hooks
Ligaments of the leg
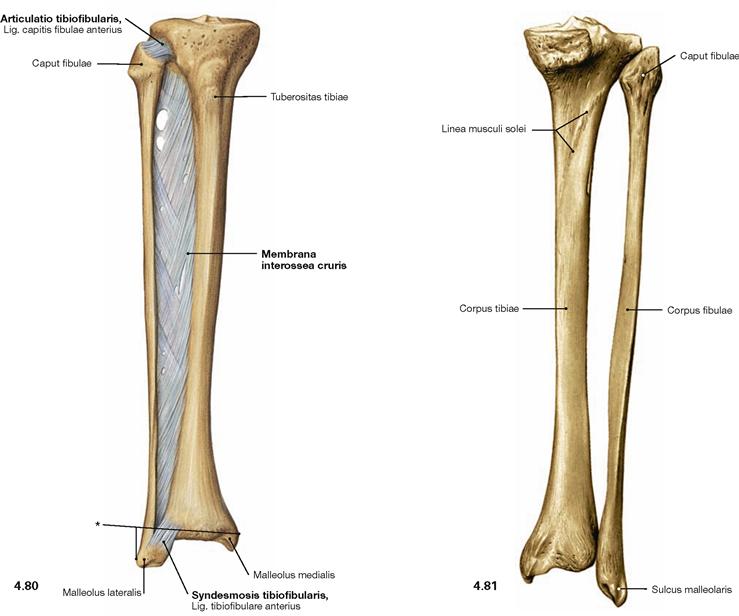
Fig. 4.80 and Fig. 4.81 Ligaments of the tibia, Tibia, and the fibula, Fibula, right side; ventral (→ Fig. 4.80) and dorsal (→ Fig. 4.81) views.
The proximal Ligg. capitis fibulae anterius and posterius create an amphiarthrosis (Articulatio tibiofibularis). Distally, both bones are fixed by the Ligg. tibiofibularia anterius and posterius in a syndesmosis (Syndesmosis tibiofibularis). Between both bones, the Membrana interossea cruris serves as an additional stabiliser with dense connective tissue and collagen fibres, which predominantly course obliquely downwards from the Tibia to the Fibula. Together with the inferior articular surface of the Tibia, the medial and lateral Malleus form the malleolar fork. The latter provides the socket for the ankle joint.
* malleolar fork
Ankle joint
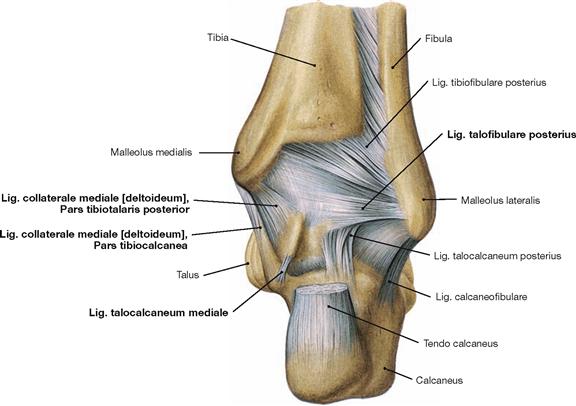
Fig. 4.83 Ankle joint (talocrural joint), Articulatio talocruralis, right side, with ligaments; dorsal view.
Parts of the Lig. collaterale mediale (Pars tibiotalaris posterior, Pars tibiocalcanea) and the lateral Lig. talofibulare posterius support the joint from the posterior side.
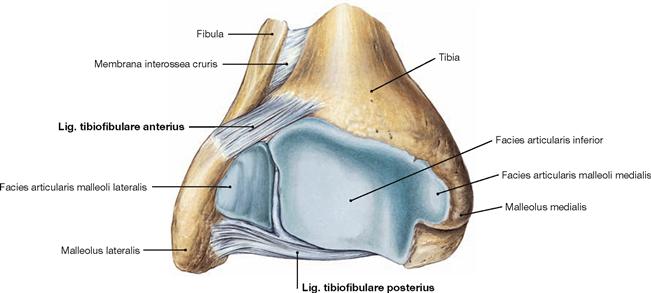
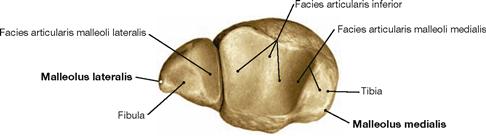
Fig. 4.84 Distal end of the tibia, Tibia, and fibula, Fibula, right side; distal view.
Tibia and Fibula are connected through the Syndesmosis tibiofibularis and together form the malleolar fork, the socket of the ankle joint.
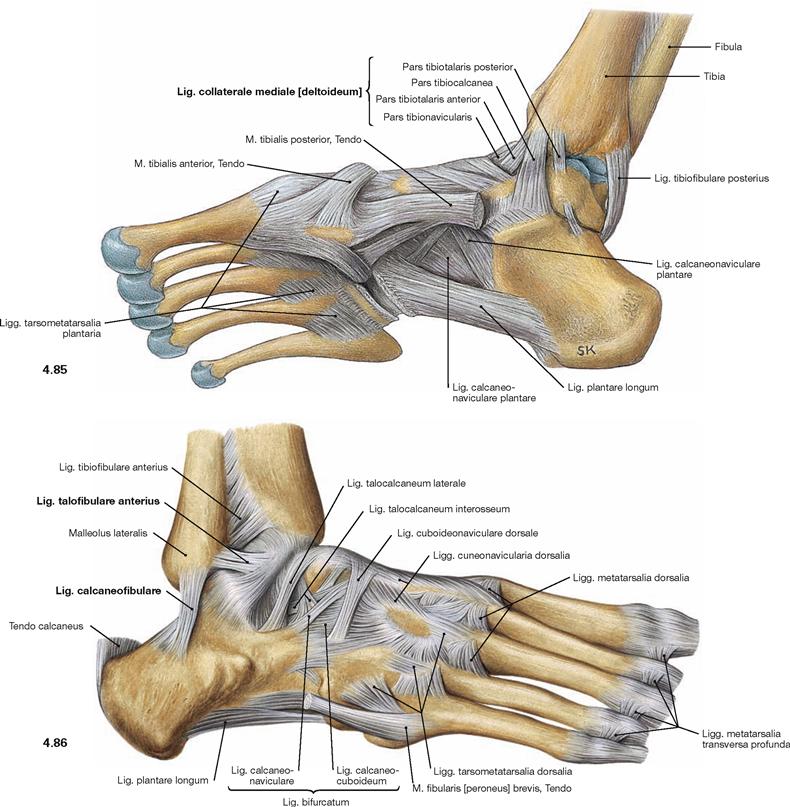
Fig. 4.85 and Fig. 4.86 Ankle joint (talocrural joint), Articulatio talocruralis, right side, with ligaments; medial (→ Fig. 4.85) and lateral (→ Fig. 4.86) views.
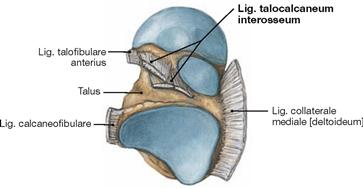
The movements of the foot take place in the (upper) ankle joint and in the (lower) talocalcaneonavicular joint. The other joints of the Tarsus and Metatarsus are amphiarthroses which increase the range of movement of the talocalcaneonavicular joint to a certain extent. In the ankle joint, the malleolar fork constitutes the socket and the trochlea of the Talus the ball of the joint. Medially, both joints are stabilised by a fan-shaped radiation of ligaments that is referred to as Lig. collaterale mediale (deltoideum) and consists of four parts (Pars tibiotalaris anterior, Pars tibiotalaris posterior, Pars tibiocalcanea, and Pars tibionavicularis) which connect the respective bones. There are three single ligaments on the lateral side (Lig. talofibulare anterius, Lig. talofibulare posterius, Lig. calcaneofibulare). These ligaments provide additional stabilisation of the talocalcaneonavicular joint.
Talocalcaneonavicular joint
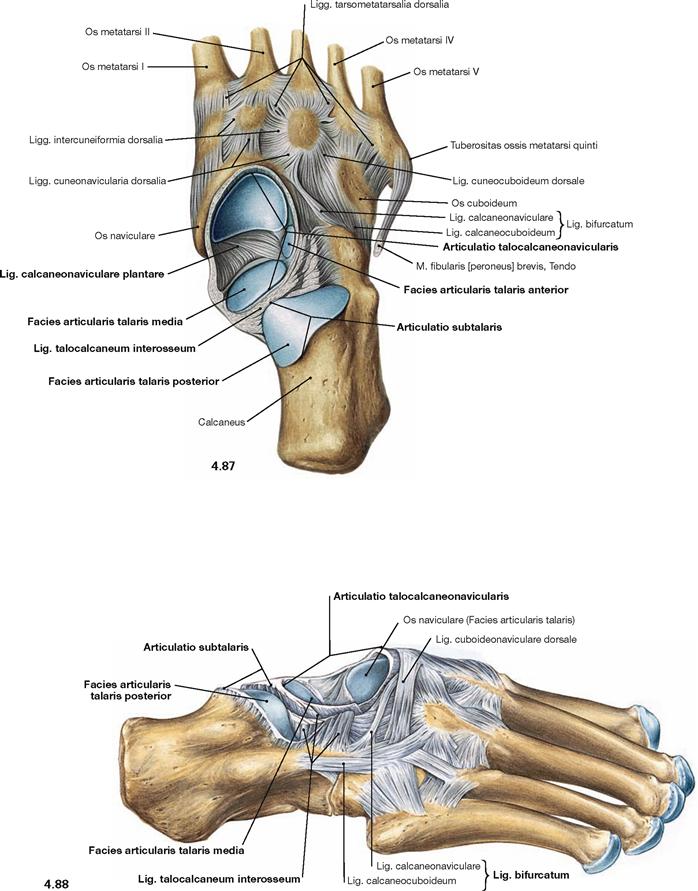
Fig. 4.87 and Fig. 4.88 Talocalcaneonavicular joint, Articulatio talocalcaneonavicularis, distal joint bodies, right side; proximal (→ Fig. 4.87) and lateral (→ Fig. 4.88) views after removal of the Talus.
In the talocalcaneonavicular joint, Talus, Calcaneus and Os naviculare articulate in two independent joints. The posterior joint (Articulatio subtalaris) is formed by the posterior corresponding articular surfaces of Talus and Calcaneus. This partial joint is separated by the Lig. talocalcaneum interosseum, positioned in the Sinus tarsi, from the anterior partial joint (Articulatio talocalcaneonavicularis). In the anterior partial joint, the anterior articular surfaces of Talus and Calcaneus articulate as well as the head of the Talus articulates with the Os naviculare anteriorly and with the Lig. calcaneonaviculare plantare inferiorly. At this contact point the latter shows an articular surface of hyaline cartilage and contributes to the plantar arch. Both parts of the joint create a functional unit and are often collectively referred to as Articulatio talocalcaneonavicularis.
In addition to the ligaments of the ankle joints, there are several ligaments which stabilize the skeletal elements of the talocalcaneonavicular joint. Besides the Lig. talocalcaneum interosseum, these are the Lig. talocalcaneum mediale and the Lig. talocalcaneum laterale (→ Figs. 4.83 und 4.86). For the range of movement in the talocalcaneonavicular joint → Figure 4.92.
Joints of the foot
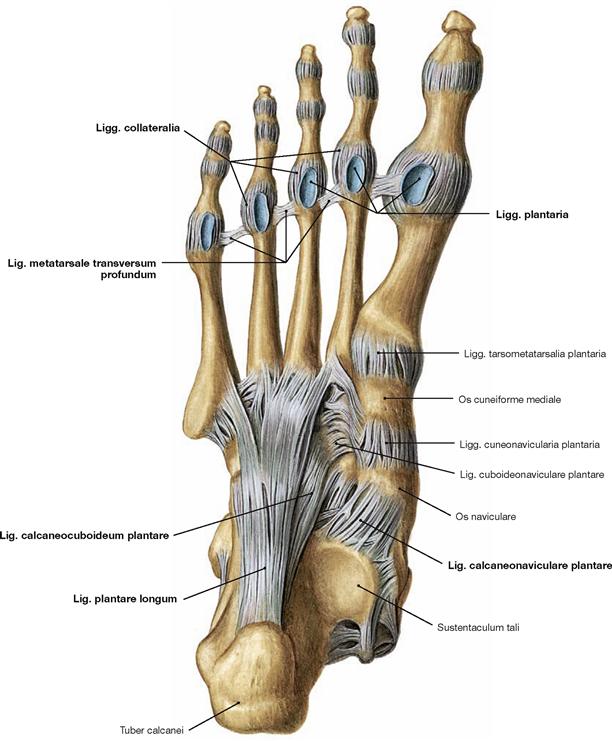
Fig. 4.90 Joints of the foot, Articulationes pedis, right side, with ligaments; plantar view.
The remaining joints of the Tarsus and Metatarsus are amphiarthroses which only minimally contribute to the movement of the foot. Together however, they extend the range of movement of the talocalcaneonavicular joint and transform the foot into an elastic base. At the Tarsus, two joints can be emphasized which contribute to supination and pronation movements of the foot. The CHOPART’s joint (Articulatio tarsi transversa) is composed of the Articulatio talonavicularis and the Articulatio calcaneocuboidea (→ Fig. 4.33). The LISFRANC’s joint (Articulationes tarsometatarsales) is the connection to the Metatarsus (→ Fig. 4.33). These two articulation lines have clinical relevance as important amputation lines.
The metatarsal bones articulate in several separate joints. The metatarsal bones are connected proximally by the Articulationes intermetatarsales and distally by the Lig. metatarsale transversum profundum. The joints of forefoot and midfoot are linked by strong plantar, dorsal, and interosseous ligaments. The CHOPART’s joint is stabilised dorsally by the Lig. bifurcatum which divides into two ligaments (Lig. calcaneonaviculare and Lig. calcaneocuboideum, → Fig. 4.87) and is opposed on the plantar side by the Lig. calcaneocuboideum plantare. Together with the Lig. calcaneonaviculare plantare, the Lig. plantare longum serves to stabilise the plantar arch. The latter is more superficial than the other plantar ligaments and spans from the Calcaneus to the Os cuboideum and the Ossa metatarsalia II–IV. The digital joints can be categorised in metatarsophalangeal joints (Articulationes metatarsophalangeales) and in proximal and distal interphalangeal joints (Articulationes interphalangeae proximales and distales). The range of movement in all digital joints is limited by tight collateral ligaments (Ligg. collateralia) and inferiorly by the Ligg. plantaria.
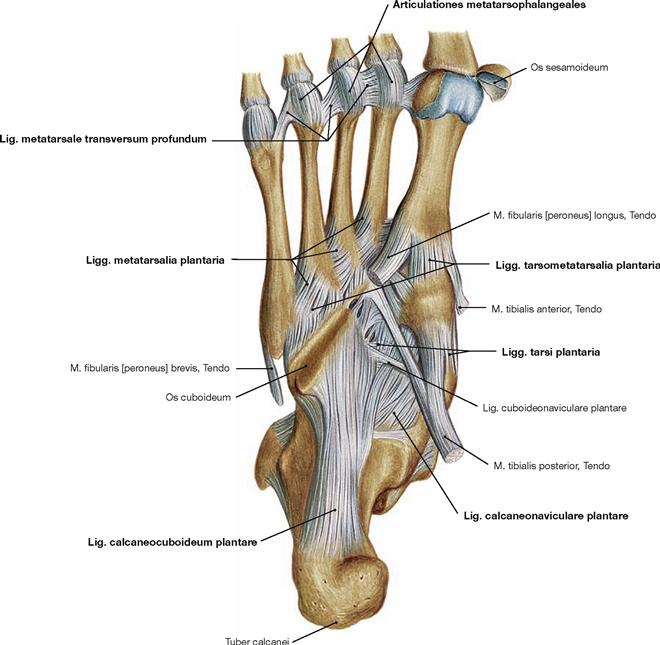
Fig. 4.91 Joints of the foot, Articulationes pedis, right side, with ligaments; plantar view; after removal of the Lig. plantare longum.
Ankle joint and other joints of the foot
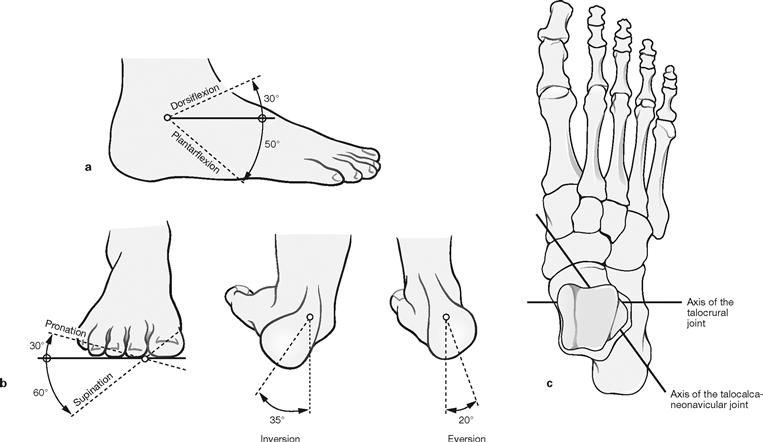
Figs. 4.92a to c Range of movement in the ankle joint and the talocalcaneonavicular joint. (according to [1])
The ankle joint is a classical hinge joint (ginglymus) allowing for dorsiflexion (extension) and plantarflexion of the foot (a). The transverse axis of the joint projects through both Malleoli (c).
The talocalcaneonavicular joint is an atypical pivot joint (Articulatio trochoidea) for which a simplified axis was defined which enters the neck of the Talus from a medial superior direction and exits the Calcaneus at the lateral posterior side (c). This joint enables inversion (sole moving towards the median plane) and eversion (sole moving outwards) of the foot. These movements of the hindfoot are complemented by the movements of the other foot joints (CHOPART’s and LISFRANC’s joint) to permit supination (lifting the medial margin of the foot) and pronation (lifting the lateral margin of the foot) (b).
Range of movement:
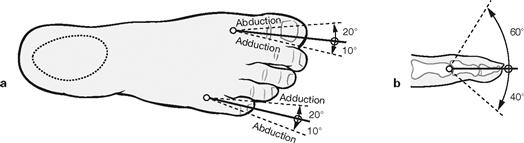
Figs. 4.93a and b Range of movement of the phalangeal joints. (according to [1])
The metatarsophalangeal joints are condyloid joints which are limited to two axes of movement by tight ligaments (rotational movements are not possible; a). The proximal and intermediate phalangeal joints are hinge joints and only allow for minimal flexion (b). More important than the active movements of the toes is their passive resistance during the rolling motion of the foot when walking.
Range of movement of the metatarsophalangeal joints:
Plantar arch
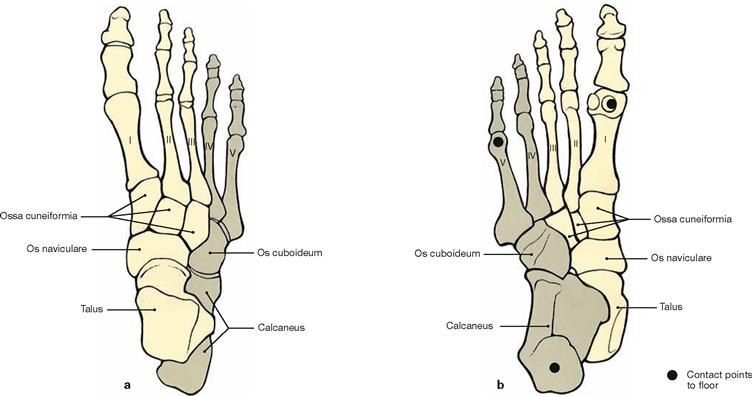
Figs. 4.94a and b Bones of the plantar arch, right side; dorsal (a) and plantar (b) views.
While the heads of the metatarsal bones are positioned in the plantar plane, the Ossa cuneiformia, Os naviculare and Talus, particularly towards their posterior aspect, position themselves on top of their lateral skeletal parts, resulting in the Talus to be placed on top of the Calcaneus. Thus, a medially open longitudinal arch is formed. The transverse arch of the foot is formed by the wedge-shaped Ossa cuneiformia and the bases of the metatarsal bones. Due to these arches, the foot has only three contact points with the floor: at the heads of the metatarsal bones I and V and at the Tuber calcanei.
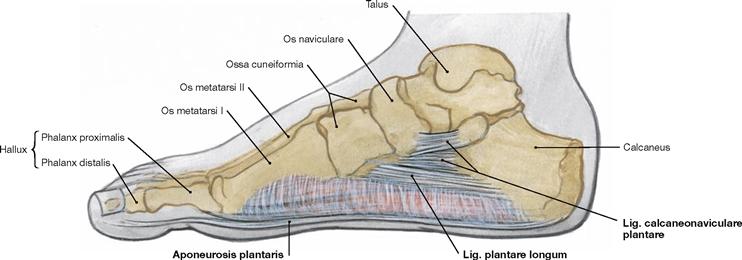
Fig. 4.95 Ligaments of the longitudinal plantar arch, right side; medial view.
The ligaments of the foot passively maintain the longitudinal arch of the foot. They are actively supported by the tendons of the M. tibialis posterior and M. fibularis longus (→ Fig. 4.148) and the short muscles on the sole of the foot. These supporting structures provide the tension band system to counteract the body weight. The ligaments can be categorized into three superimposing levels:
Imaging
Pelvis
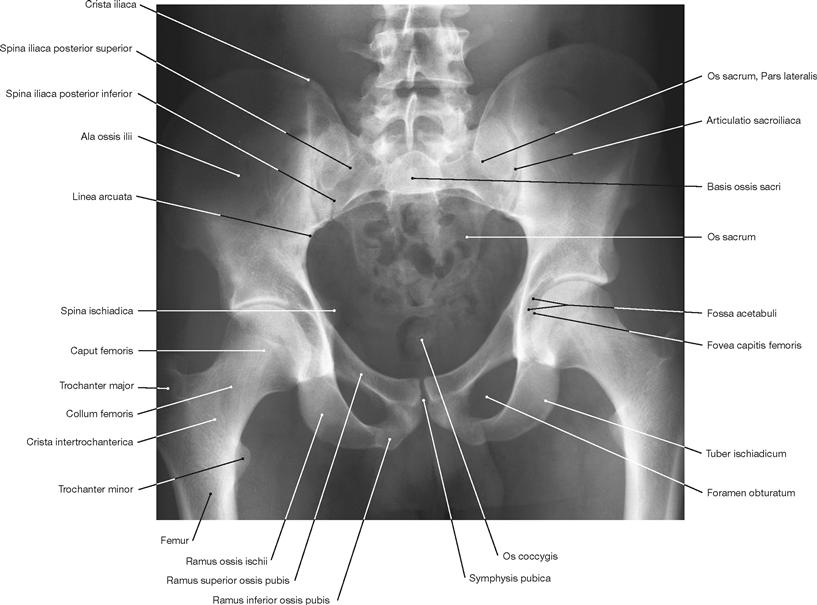
Fig. 4.96 Pelvis of a man; radiograph in anteroposterior (AP) beam projection; upright standing position.
Hip joint
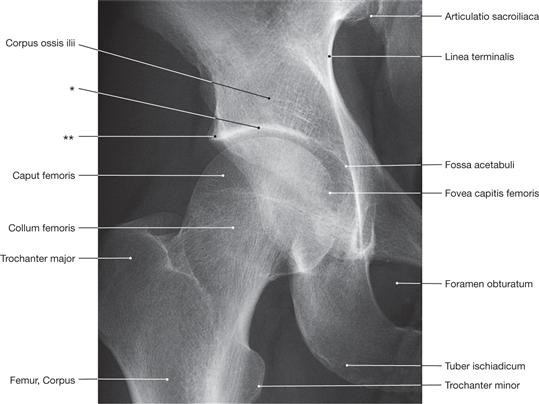
Fig. 4.97 Hip joint, Articulatio coxae, right side; radiograph in anteroposterior (AP) beam projection; upright standing position.
* clinical term: roof of the acetabulum
** clinical term: notch at the roof of the acetabulum
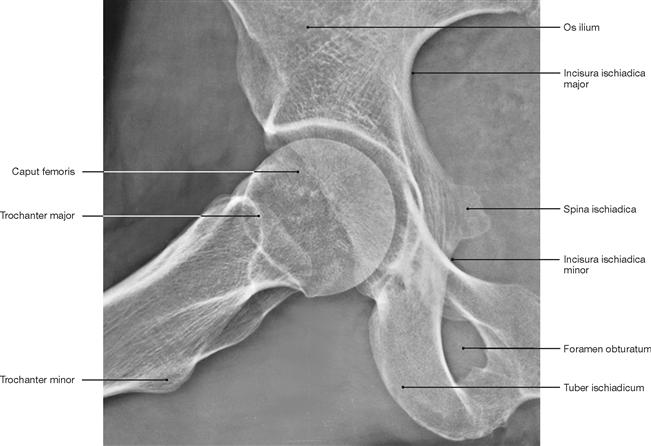
Fig. 4.98 Hip joint, Articulatio coxae, right side; radiograph in LAUENSTEIN projection (abduction and flexion of the thigh in supine position).
Knee joint
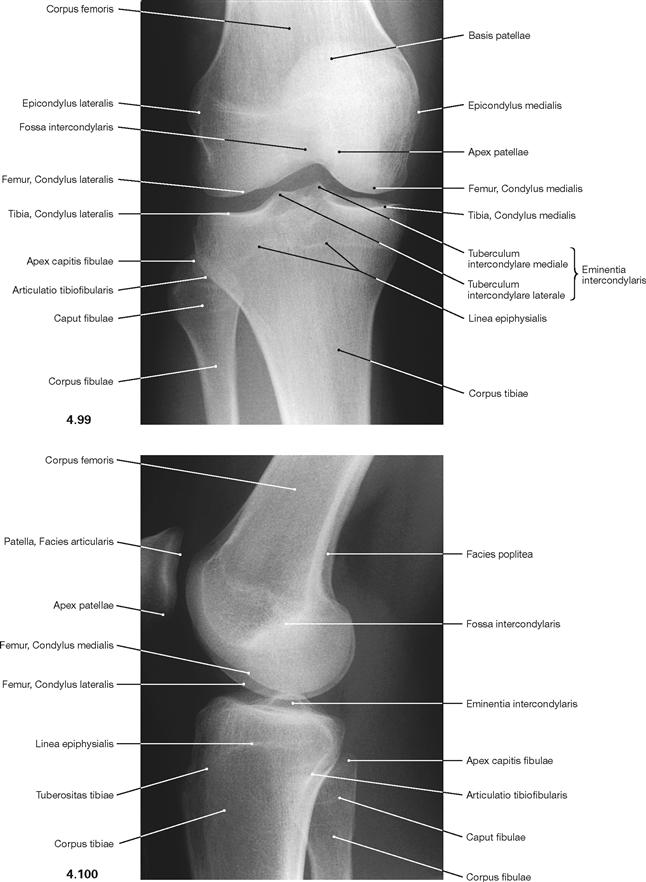
Fig. 4.99 and Fig. 4.100 Knee joint, Articulatio genus, radiograph in anteroposterior (AP) beam projection (→ Fig. 4.99) and in lateral beam projection (→ Fig. 4.100); in supine position.
It has to be considered that the contours of the medial and lateral femoral condyles are not congruent.
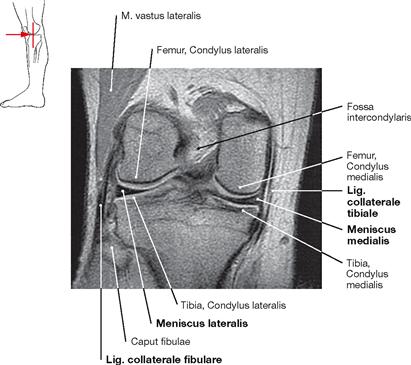
Fig. 4.101 Knee joint, Articulatio genus, right side; magnetic resonance imaging (MRI) sagittal section; ventral view.
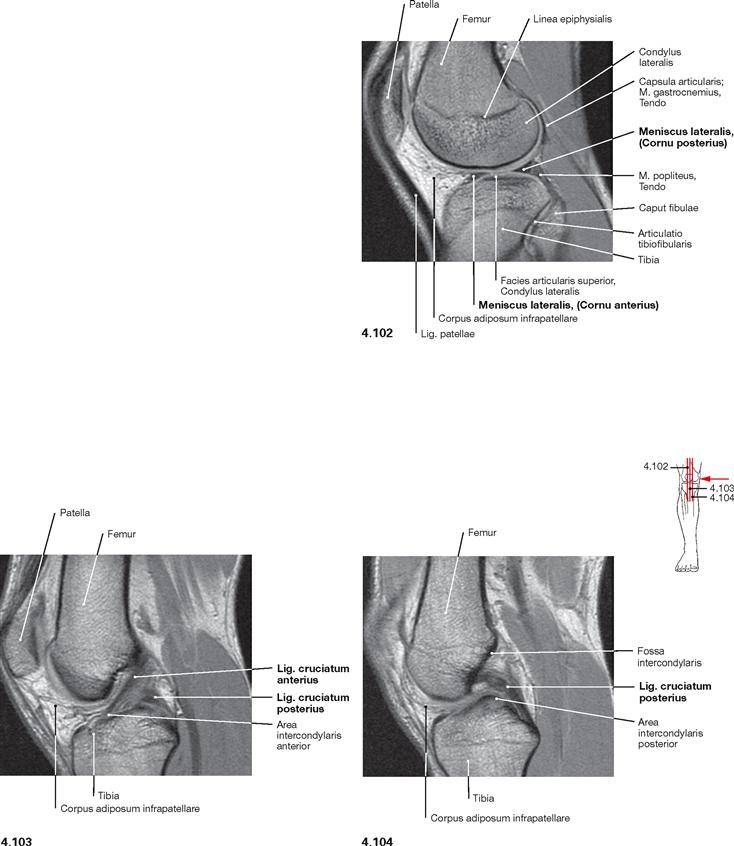
Fig. 4.102 to Fig. 4.104 Knee joint, Articulatio genus, right side; magnetic resonance imaging (MRI) sagittal sections; medial view.
Compact bone appears dark with this imaging technique.
Ankle joint and talocalcaneonavicular joint
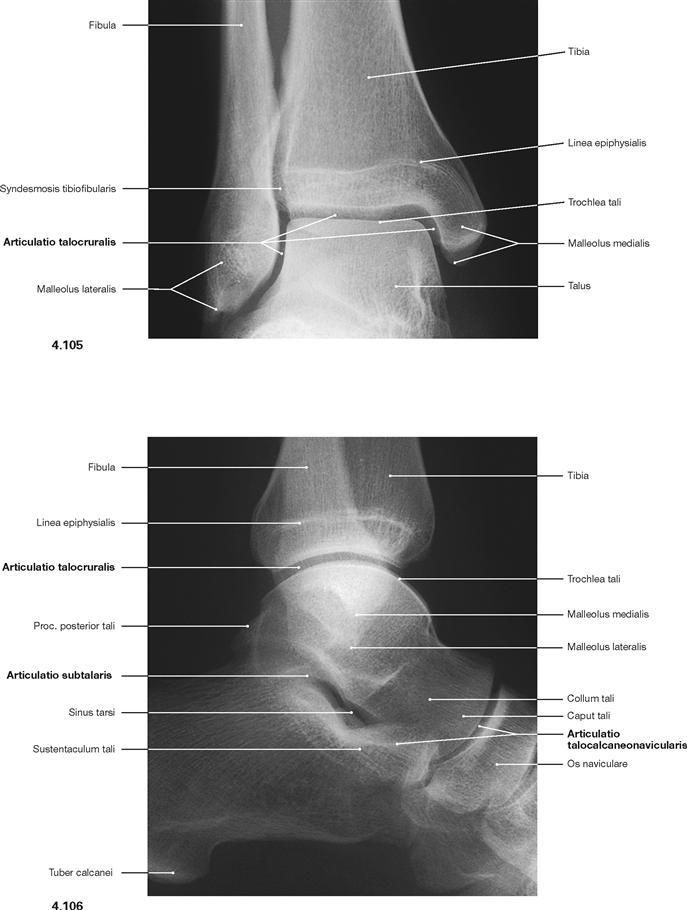
Fig. 4.105 and Fig. 4.106 Ankle joint (talocrural joint), and talocalcaneonavicular joint, Articulationes talocruralis and talocalcaneonavicularis, right side; radiograph in anteroposterior (AP) beam projection (→ Fig. 4.105), and in lateral beam projection (→ Fig. 4.106).
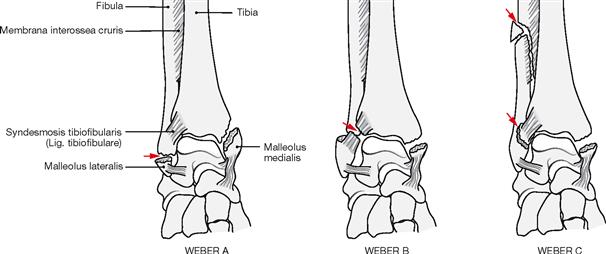
Fractures of the ankle joint
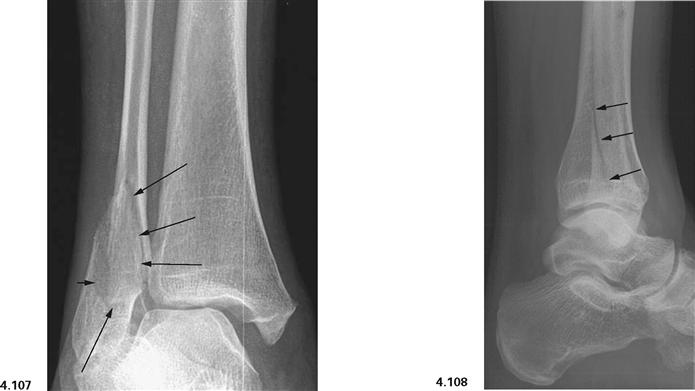
Fig. 4.107 and Fig. 4.108 Ankle joint (talocrural joint), Articulatio talocruralis, right side, with malleolar fracture (WEBER type B); radiograph in anteroposterior (AP) beam projection (→ Fig. 4.107), and in lateral beam projection (→ Fig. 4.108). [17]
Fracture lines are marked with arrows.
Fascias of the lower extremity
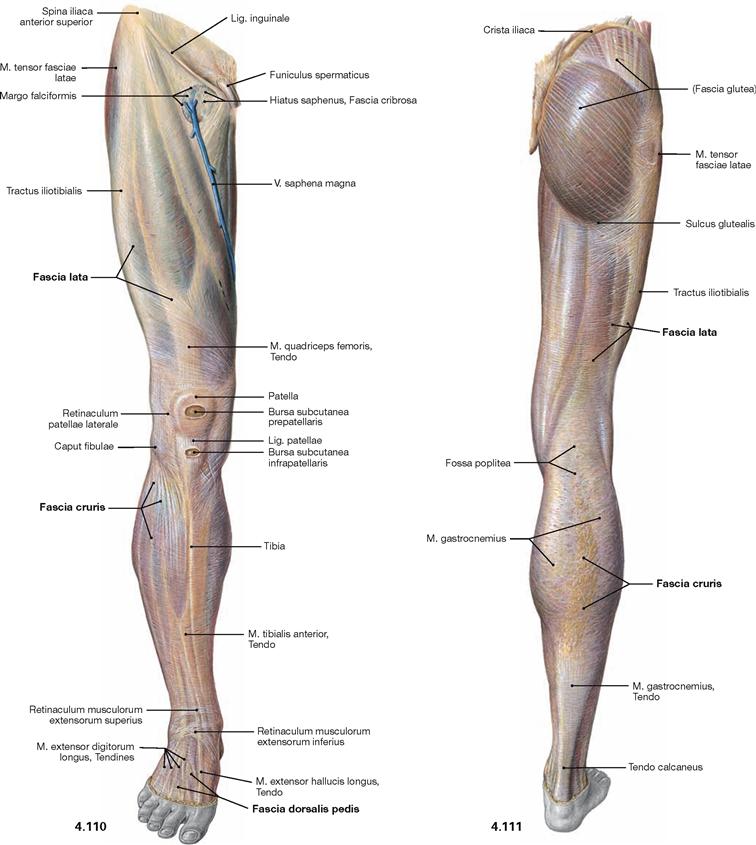
Fig. 4.110 and Fig. 4.111 Fasciae of the thigh, Fascia lata, the leg, Fascia cruris, and the dorsum of the foot, Fascia dorsalis pedis, right side; ventral (→ Fig. 4.110) and dorsal (→ Fig. 4.111) views.
Muscles
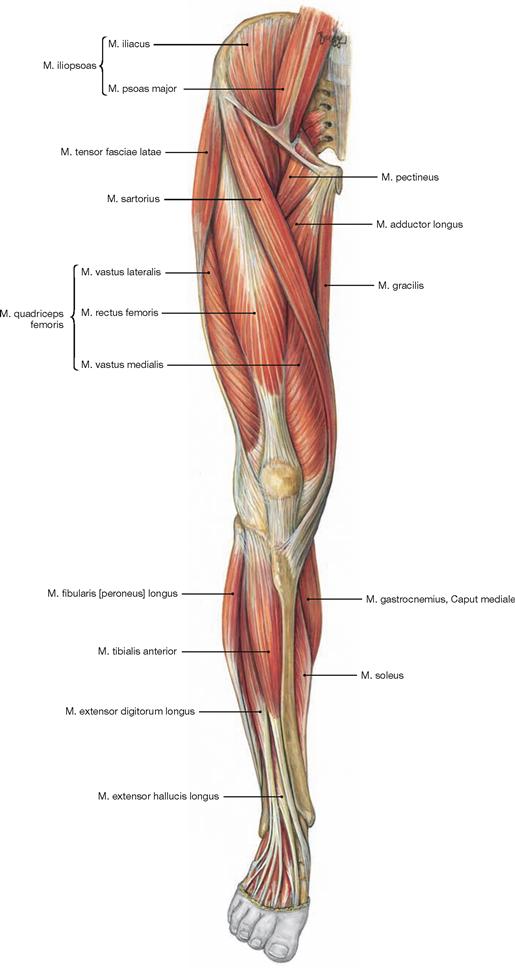


Muscles of the hip and thigh
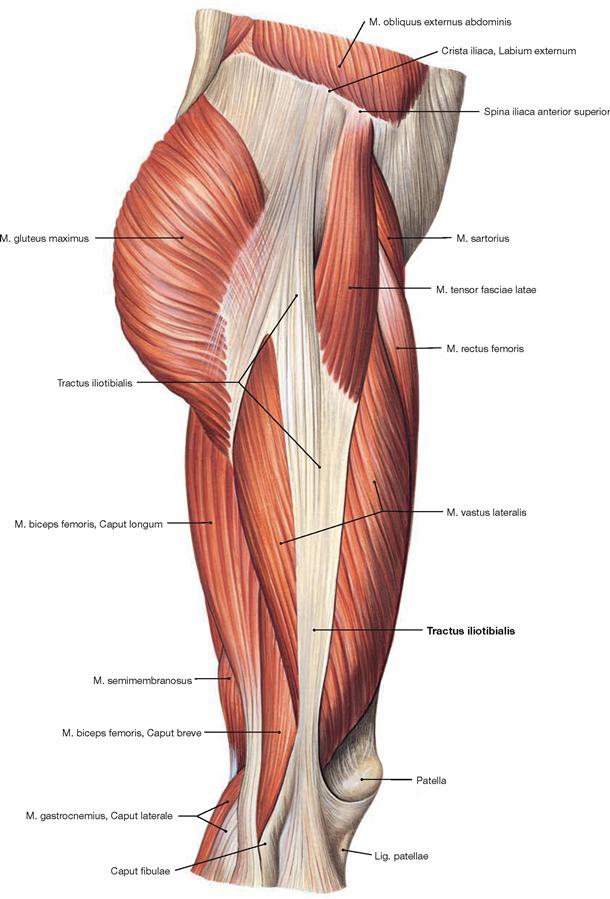
Fig. 4.114 Muscles of the hip and thigh, right side; lateral view.
The Tractus iliotibialis serves as reinforcement of the fascia of the thigh (Fascia lata) and connects the ilium with the Tibia. It counterbalances the body weight-induced medial forces on the thigh bone. This principle is referred to as tension band effect.
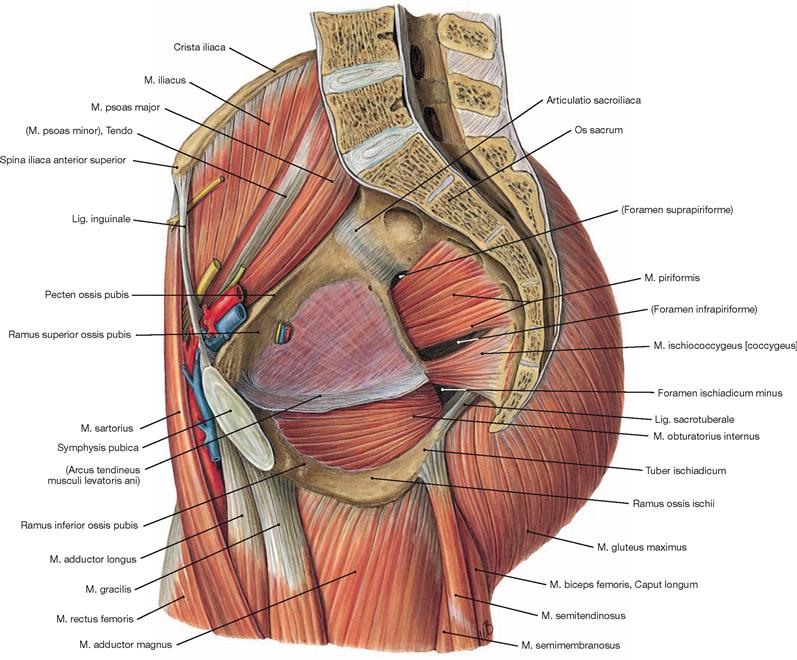

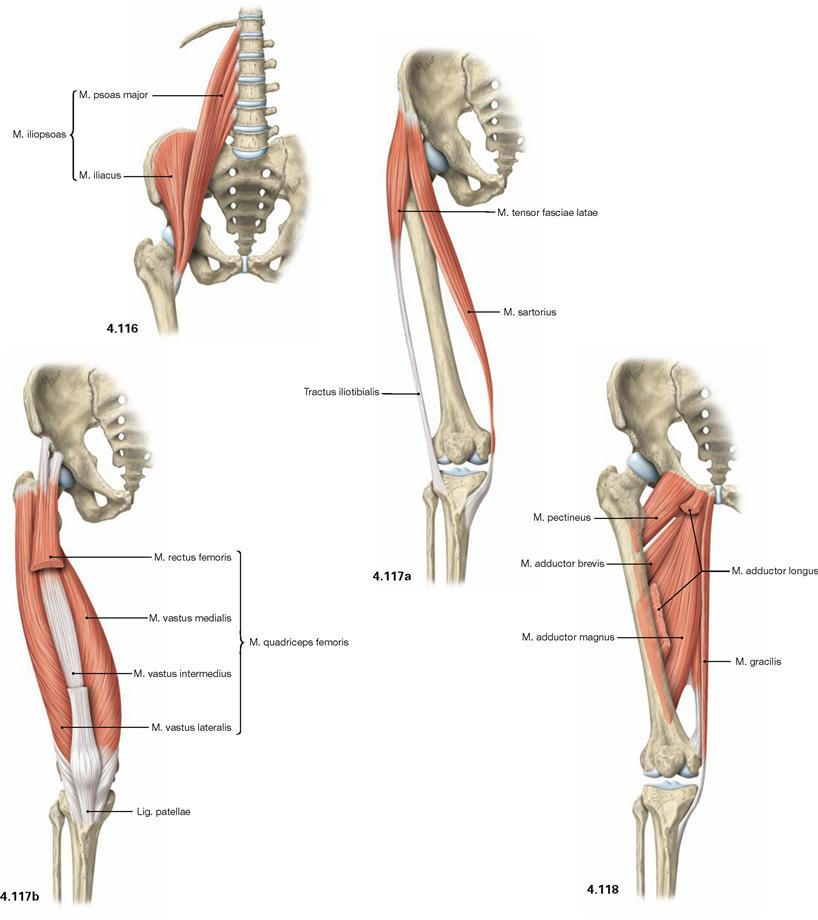
Fig. 4.116 to Fig. 4.118 Ventral muscles of the hip and thigh and medial muscles of the thigh, right side; ventral view.
The muscles of the hip and thigh are equally important to erect the body from the supine position, to maintain an upright position, and for the normal gait. The ventral muscles of the hip comprise the M. iliopsoas (→ Fig. 4.116) which functions as most important flexor of the hip. Located at the lateral thigh, the M. tensor fasciae latae (→ Fig. 4.117a) functions as tension band via its insertion on the iliotibial tract and protects the thigh bone from fractures by reducing bending stress. Together with the M. sartorius (→ Fig. 4.117a), the M. tensor fasciae latae flexes the hip joint. Due to its innervation, the M. tensor fasciae latae is also counted among the dorsolateral hip muscles.
The four-headed M. quadriceps femoris (→ Fig. 4.117b) is the only extensor of the knee joint and is essential to erect the body from a squatting position. Its M. rectus femoris spans two joints and also flexes the hip.
Located medially, the muscles of the adductor group (Mm. adductores, → Fig. 4.118) are the most important adductors of the thigh and stabilise the hip during standing and walking.
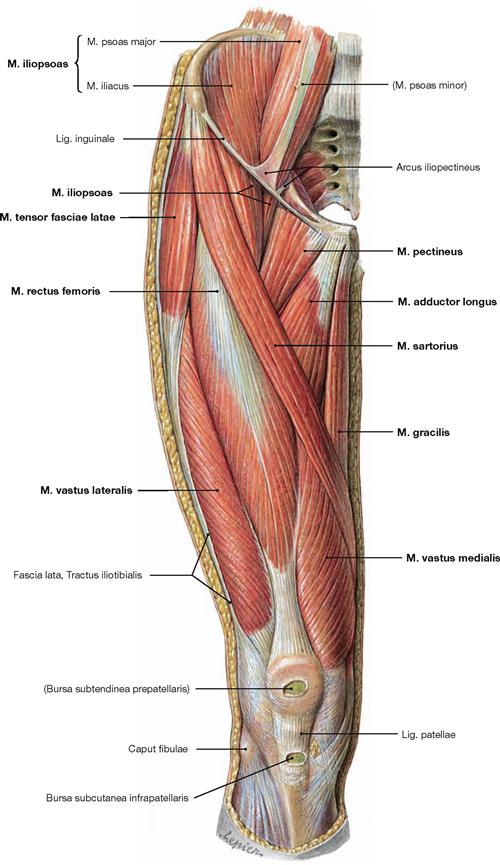
Fig. 4.119 Ventral muscles of the hip and thigh, and medial muscles of the thigh, right side; ventral view; after removal of the Fascia lata ventral to the Tractus iliotibialis.
The M. iliopsoas is composed of two different muscles which originate from the lumbar part of the vertebral column (M. psoas) and the Fossa iliaca (M. iliacus). Inferior to the inguinal ligament, only a short portion of both parts of the muscle courses to the common insertion site at the Trochanter minus.
The M. sartorius is ensheathed by a split portion of the Fascia lata and crosses the anterior aspect of the thigh to insert at the medial aspect of the Tibia posterior to the transverse axis of the knee. Thus, it flexes the hip and the knee.
Medially, the muscles of the adductor group are located on top of each other in several layers of which only the superficial M. pectineus, M. adductor longus, and M. gracilis are visible. The four heads of the M. quadriceps femoris (M. rectus femoris, Mm. vasti lateralis, medialis, and intermedius) lie distally and laterally of the M. sartorius. Their common tendon incorporates the Patella as a sesamoid bone before the fibres continue as Lig. patellae to the Tuberositas tibiae.
Most laterally, the M. tensor fasciae latae inserts in the Tractus iliotibialis. The common insertion of the Mm. sartorius, gracilis, and semitendinosus inferior to the medial tibial condyle is often referred to as the “Pes anserinus superficialis”.
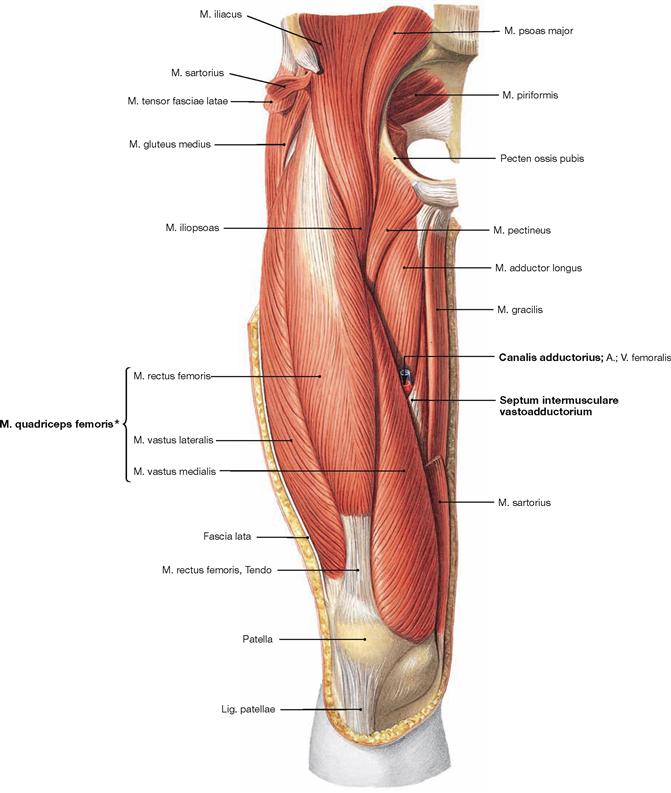
Fig. 4.120 Ventral muscles of the hip and thigh, and medial muscles of the thigh, right side; ventral view; after removal of the Fascia lata, the M. sartorius, and the M. tensor fasciae latae.
After removal of the M. sartorius, the entrance to the adductor canal (Canalis adductorius) is visible which is demarcated dorsally by the M. adductor longus. In its anterior portion, the canal is covered by the Septum intermusculare adductorium which connects the fasciae of the M. vastus medialis, Mm. adductores longus and magnus.
The four heads of the M. quadriceps femoris (M. rectus femoris, Mm. vasti lateralis, medialis and intermedius) are located laterally to the adductor canal.
* The fourth head of the M. quadriceps femoris, the M. vastus intermedius, lies beneath the M. rectus femoris.
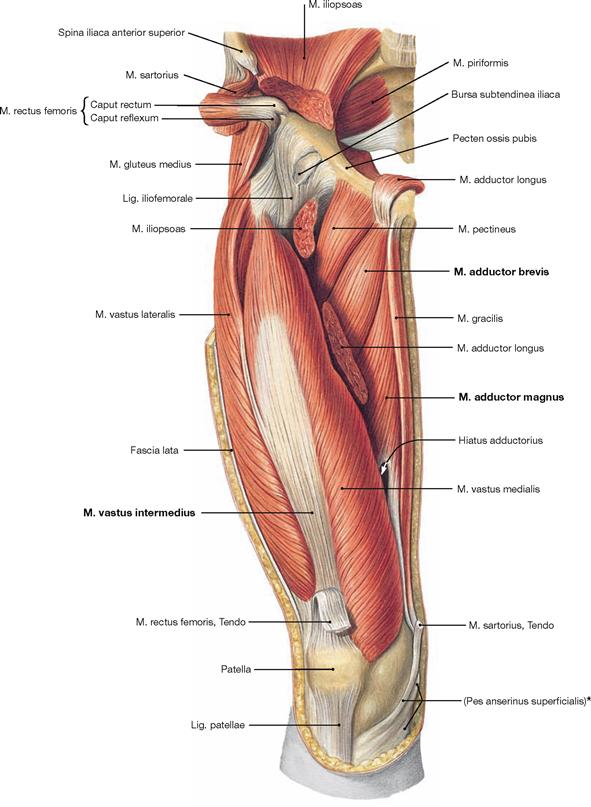
Fig. 4.121 Ventral muscles of the hip and thigh, and deep medial muscles of the thigh, right side; ventral view; after removal of the Fascia lata, Mm. sartorius, rectus femoris, and adductor longus, and partial removal of the M. iliopsoas at the area of the hip joint.
The M. rectus femoris and a part of the M. adductor longus are reflected superiorly. After removal of the M. rectus femoris, the M. vastus intermedius of the M. quadriceps femoris is visible. The resection of the M. sartorius and M. adductor longus reveals the deep adductor muscles, the M. adductor brevis and parts of the M. adductor magnus.
* common insertion of the Mm. sartorius, gracilis and semitendinosus
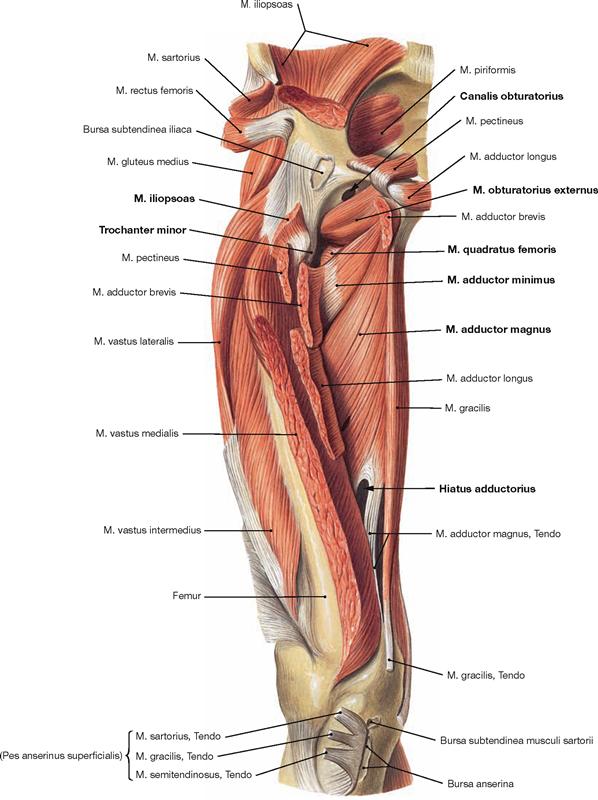
Fig. 4.122 Ventral muscles of the hip and thigh, and deep medial muscles of the thigh, right side; ventral view; after almost complete resection of the superficial and some of the deep muscles.
Upon reflecting the superficial adductor muscles and the M. adductor brevis laterally, the M. adductor magnus becomes visible. Its upper portion is also referred to as M. adductor minimus. The M. adductor magnus and its tendon form the adductor hiatus (Hiatus adductorius) through which the blood vessels of the thigh (A./V. femoralis) pass to reach the popliteal fossa. Proximal, the insertion of the M. iliopsoas at the Trochanter minor is recognisable after resection of the M. pectineus and M. adductor brevis. The Canalis obturatorius is displayed as opening within the Membrana obturatoria. It serves as neurovascular passageway between the small pelvis and the thigh. Caudal of this opening, the almost horizontal fibres of the M. obturatorius externus and the M. quadratus femoris are revealed, both of which belong to the pelvitrochanteric group of dorsal hip muscles (→ p. 306). These muscles are often not displayed during the dissection classes and thus, their classes is more difficult to envision.
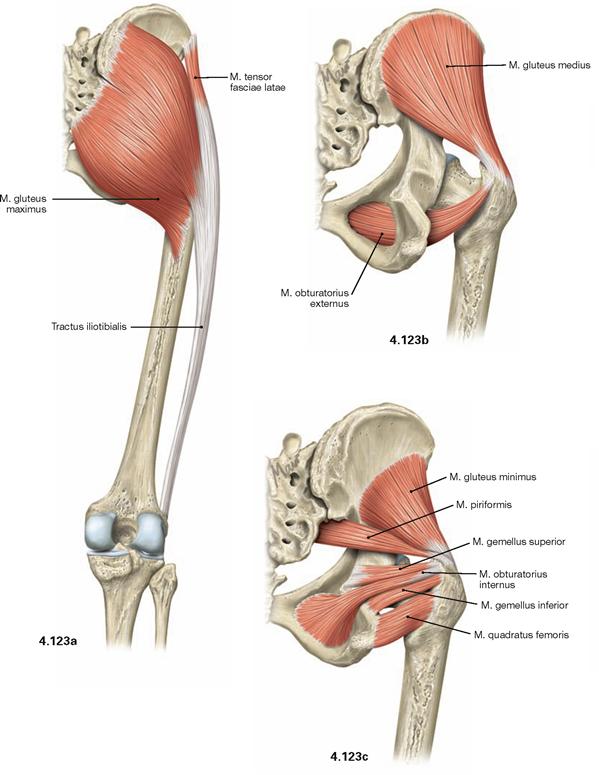
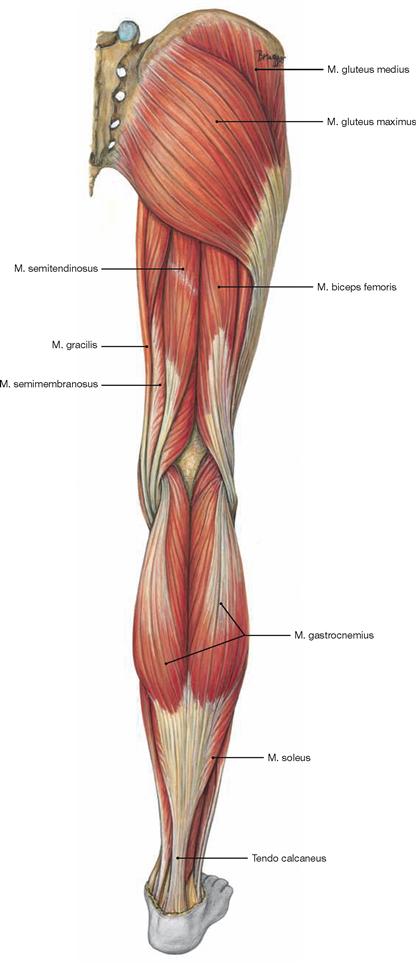
Figs. 4.123a to c Dorsal muscles of the hip, right side; dorsal view.
The dorsal muscles of the hip are categorized in a dorsolateral and a pelvitrochanteric group.
The dorsolateral group comprises the Mm. glutei maximus, medius and minimus. According to its innervation, the M. tensor fasciae latae (→ Fig. 4.117a) also may be counted among this group. The M. gluteus maximus (→ Fig. 4.123a) is the most important extensor and lateral rotator of the hip and for example necessary when climbing stairs. In contrast, the smaller gluteal muscles (Mm. glutei medius and minimus, → Figs. 4.123b and c) are the most important abductors and medial rotators of the thigh. Their action stabilises the hip during standing and walking and prevents the tilting of the pelvis to the contralateral side when standing on one leg (for the function of the small gluteal muscles and the TRENDELENBURG’s sign → p. 335).
The pelvitrochanteric group (M. priformis, Mm. obturatorii internus and externus, Mm. gemelli superior and inferior, M. quadratus femoris → Fig. 4.123c) comprises exclusively lateral rotators.
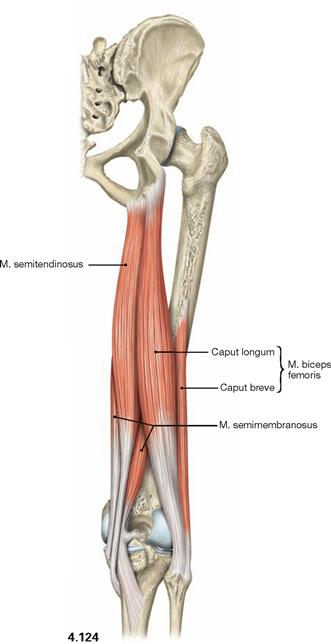
Fig. 4.124 Dorsal (ischiocrural, hamstring) muscles of the thigh, right side; dorsal view.
The dorsal (ischiocrural, hamstring) muscles (→ Fig. 4.124) on the posterior side of the thigh originate from the Tuber ischiadicum and insert to both bones of the lower leg. These muscles span two joints and facilitate extension in the hip joint while serving as strongest flexors in the knee joint. In addition, the lateral M. biceps femoris functions in lateral rotation on both joints, whereas the medial M. semitendinosus and M. semimembranosus function in medial rotation.
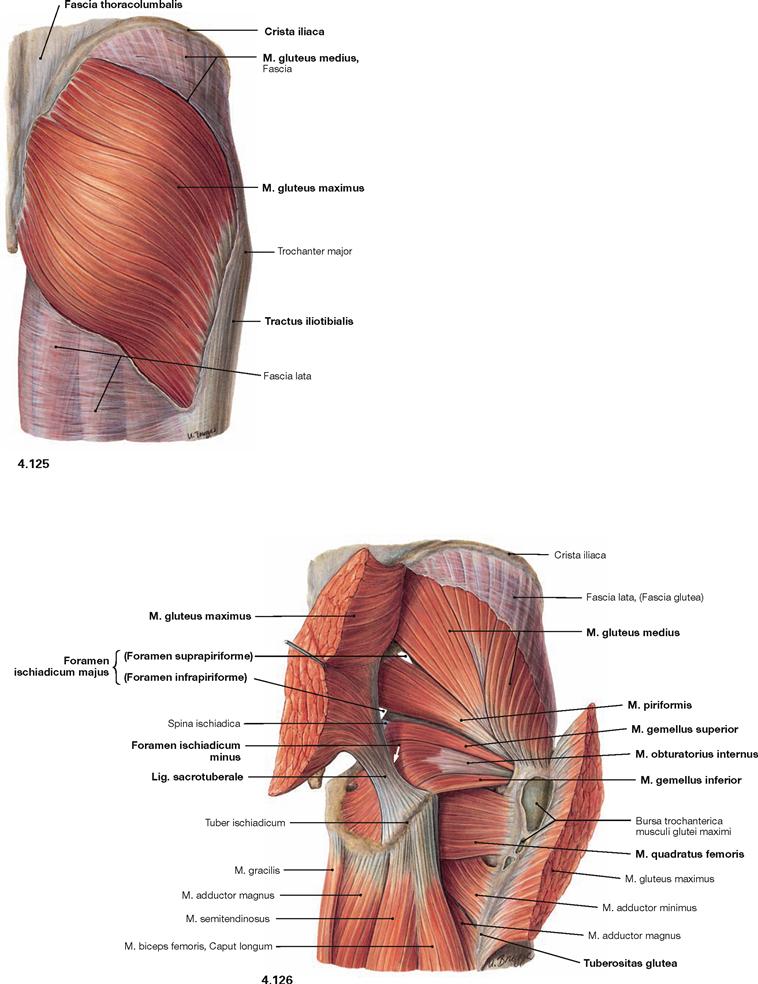
Fig. 4.125 and Fig. 4.126 Dorsal muscles of the hip and thigh, right side; dorsal view; after splitting of the Fascia lata (→ Fig. 4.125) and separation of the M. gluteus maximus (→ Fig. 4.126).
The illustration shows the superficial and the deep origins and insertions of the M. gluteus maximus. Superficially, the muscle originates from the posterior side of the sacrum, the Crista iliaca and the Fascia thoracolumbalis as well as deeply from the Lig sacrotuberale. Its muscle fibres course in an oblique way, whereas the M. gluteus medius beneath has an almost vertical orientation. The M. gluteus maximus has superficial insertions at the Fascia lata and the Tractus iliotibialis and deep insertions on the Tuberositas glutea of the Femur. Separation and lateral reflexion of the M. gluteus maximus reveals the other parts of the M. gluteus medius and the pelvitrochanteric muscles.
The M. piriformis divides the Foramen ischiadicum majus into the Foramina suprapiriforme and infrapiriforme which serve as important passageways for neurovascular structures from the pelvis. It should be noted that the M. obturatorius internus frequently continues as a tendinous structure from its deflecting point (hypomochlion) at the Incisura ischiadica minor to its insertion at the Fossa trochanterica.
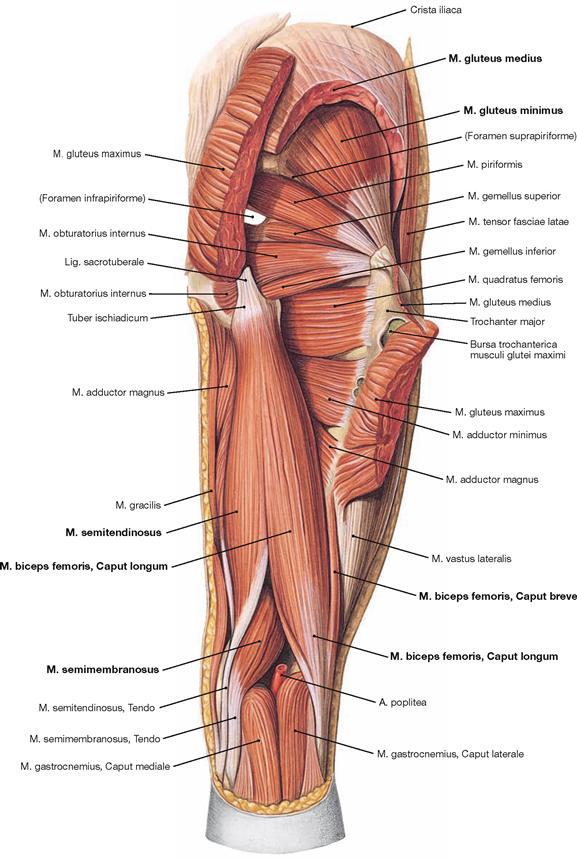
Fig. 4.127 Dorsal muscles of the hip and thigh, right side; dorsal view; after partial resection of the Mm. glutei maximus and medius.
After cutting the M. gluteus medius in addition to the M. gluteus maximus, the M. gluteus minimus is visible. Collectively, the Mm. glutei medius and minimus are referred to as small gluteal muscles. Both muscles serve for hip abduction and stabilisation of the pelvis during one-leg stand.
The dorsal side of the thigh contains the hamstring muscles which span from the Tuber ischiadicum to the bones of the lower leg. Located medially is the M. semitendinosus (named after its long tendon) and beneath the M. semimembranosus (named after its flat tendon); positioned laterally is the M. biceps femoris. The Caput longum of the latter originates from the Tuber ischiadicum, whereas the Caput breve originates from the distal thigh (Labium laterale of the Linea aspera).
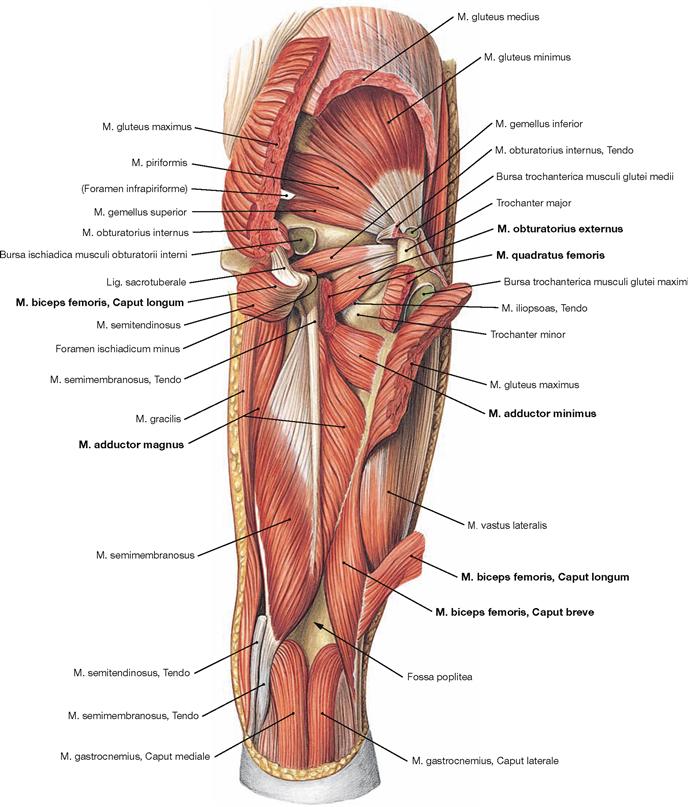
Fig. 4.128 Deep dorsal muscles of the hip and thigh, right side; dorsal view; after almost complete resection of the superficial gluteal and hamstring muscles.
Upon splitting the M. quadratus femoris, the deeper M. obturatorius externus is visible; its course is often difficult to imagine. Removal of the long head of the M. biceps femoris exposes the deep components of the adductor group. The M. adductor magnus has two functionally independent muscle parts with distinct innervation. Its major component originates from the inferior pubic ramus (this part is sometimes referred to as M. adductor minimus) and the ischial ramus. The posterior part derives from the Tuber ischiadicum and, according to its function and innervation, is counted among the hamstring muscles.
Muscles of the thigh
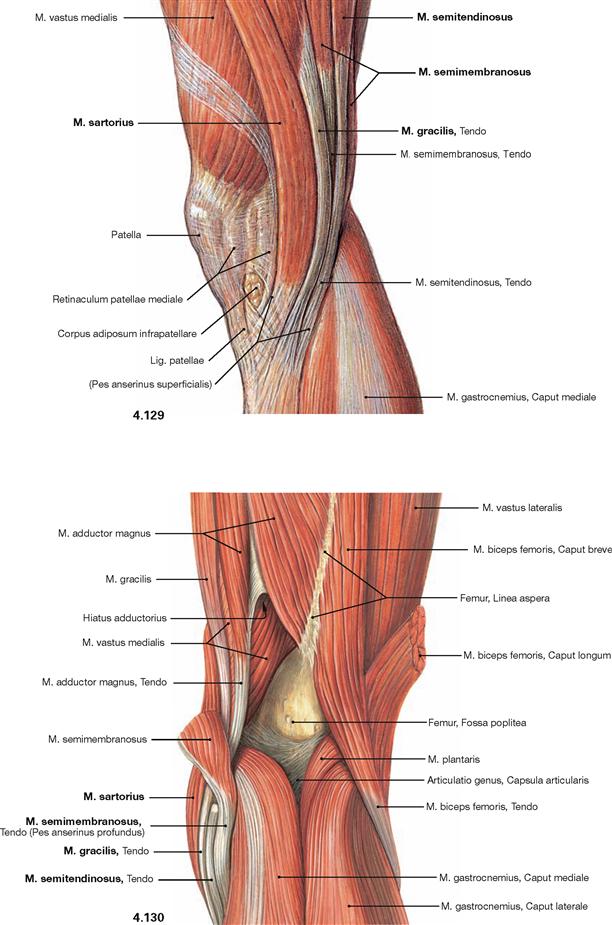
Fig. 4.129 and Fig. 4.130 Muscles in the region of the knee joint, right side; medial (→ Fig. 4.129) and dorsal (→ Fig. 4.130) views.
The common insertion of the Mm. sartorius, gracilis, and semitendinosus beneath the medial condyle of the Tibia is referred to as “Pes anserinus superficialis”. The deeply located insertion of the M. semimembranosus is called “Pes anserinus profundus”.
Muscles of the leg
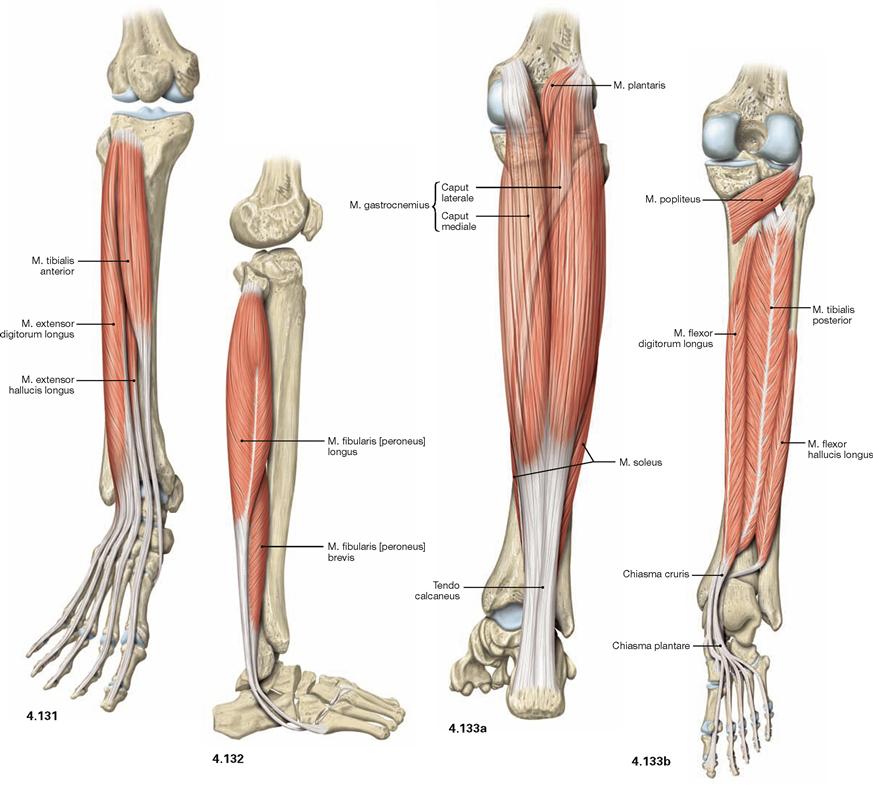
Fig. 4.131 to Fig. 4.133 Muscles of the leg, right side; ventral (→ Fig. 4.131), lateral (→ Fig. 4.132), and dorsal (→ Fig. 4.133) views.
The leg has three muscle groups. To understand their function, the position in relation to the axes of movement in the joints of the ankle and foot are important. All muscles coursing anterior to the transverse axis of the ankle joint are extensors (dorsiflexors), all muscles dorsal to this axis are flexors (plantarflexors) of the foot. All muscles with tendons coursing medial to the oblique axis of the talocalcaneonavicular joint function as supinators and lift the medial margin of the foot. Muscles with tendons lateral to this axis lift the lateral margin of the foot and thus perform pronation.
The ventral muscles of the leg function as extensors (→ Fig. 4.131). They extend the ankle joint and the talocalcaneonavicular joint, together with the other joints of the foot, they mainly support pronation. The M. tibialis anterior is the most important extensor (→ Fig. 4.131), whereas the M. extensor digitorum longus and M. extensor hallucis longus also extend the toes.
The lateral (fibular) muscles of the leg (→ Fig. 4.132) comprise the Mm. fibularis longus and brevis. They are the most important pronators and function as plantarflexors in the ankle joint due to their tendons positioned behind the flexion-extension axis. Dorsally located are the true flexor muscles (plantarflexors) which can be divided in a superficial and a deep group.
The M. triceps surae (→ Fig. 4.133a) is part of the superficial dorsal muscles and comprises the two-headed M. gastrocnemius and the M. soleus beneath. The M. triceps surae is the strongest flexor and major supinator of the foot. The M. plantaris is rather insignificant.
The deep dorsal muscles (flexors; → Fig. 4.133b) are largely equivalent to the extensors on the ventral side. The M. tibialis posterior is a flexor and a strong supinator. The M. flexor digitorum longus and M. flexor hallucis longus flex the phalangeal joints. A special role has the M. popliteus which stabilises the knee joint. Above the medial Malleolus, the tendon of the M. flexor digitorum crosses the tendon of the M. tibialis posterior (Chiasma cruris) and at the level of the sole of the foot, it crosses the tendon of the M. flexor hallucis longus (Chiasma plantare).
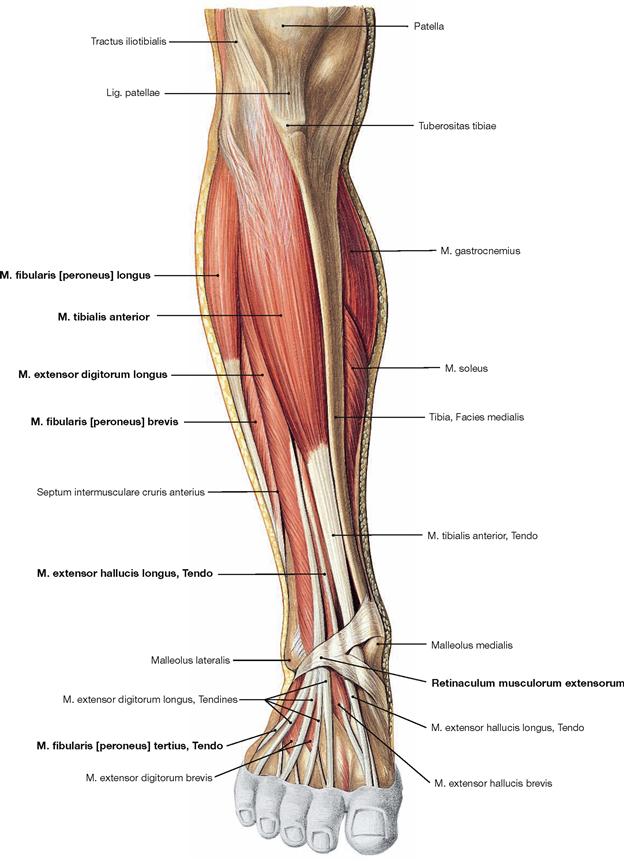
Fig. 4.134 Ventral and lateral muscles of the leg and the foot, right side; ventral view.
The M. tibialis anterior of the extensor group can be palpated near the margin of the Tibia. Since its tendon courses medial to the axis of the talocalcaneonavicular joint, it functions as a (although weak) supinator in contrast to the other extensors. The M. extensor digitorum longus derives from the proximal Tibia and Fibula, and the M. extensor hallucis longus is located between the other two extensors at the distal leg. Occasionally, the M. extensor digitorum longus shows a separation that inserts at the Os metatarsi V and is confusingly called M. fibularis tertius. In the distal part, the tendons are guided by a reinforcement of the fascia of the leg, the Retinaculum musculorum extensorum. The retinacula of the foot function as retainers and prevent the tendons from lifting off the bones during extension of the foot. Both muscles of the fibularis group (Mm. fibulares longus and brevis) belong to the lateral group and originate from the proximal and distal Fibula. Clinically, they are often referred to by their old name as peroneal muscles (fibula, greek: perone).
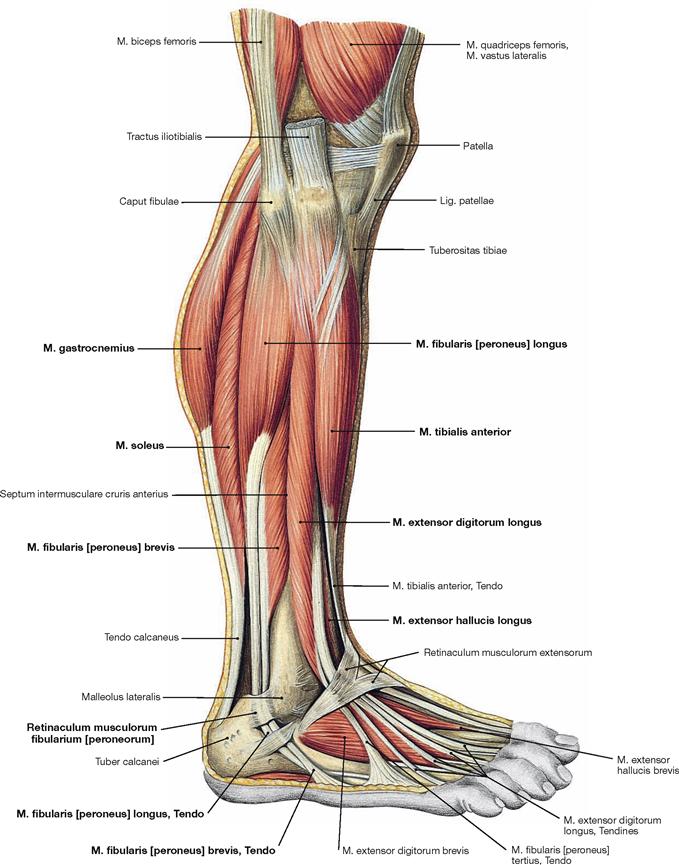
Fig. 4.135 Muscles of the leg and the foot, right side; lateral view.
In the lateral view, all three muscle groups of the leg are visible. Laterally behind the anterior group of extensors lie the fibularis muscles, dorsally lie the flexors. Since the deep flexors of the rear side are directly adjacent to the bones of the leg, only the superficial muscles (M. triceps surae), the M. gastrocnemius and the M. soleus, can be seen. The tendons of the fibularis group are guided by the Retinacula musculorum fibularium. The M. fibularis brevis inserts at the Os metatarsi V, whereas the tendon of the M. fibularis longus extends beneath the sole of the foot and inserts at the Os metatarsi I and Os cuneiforme mediale, thus actively supporting the plantar arch. It should be noted that the M. extensor hallucis longus is found distally between the M. tibialis anterior and the M. extensor digitorum longus.
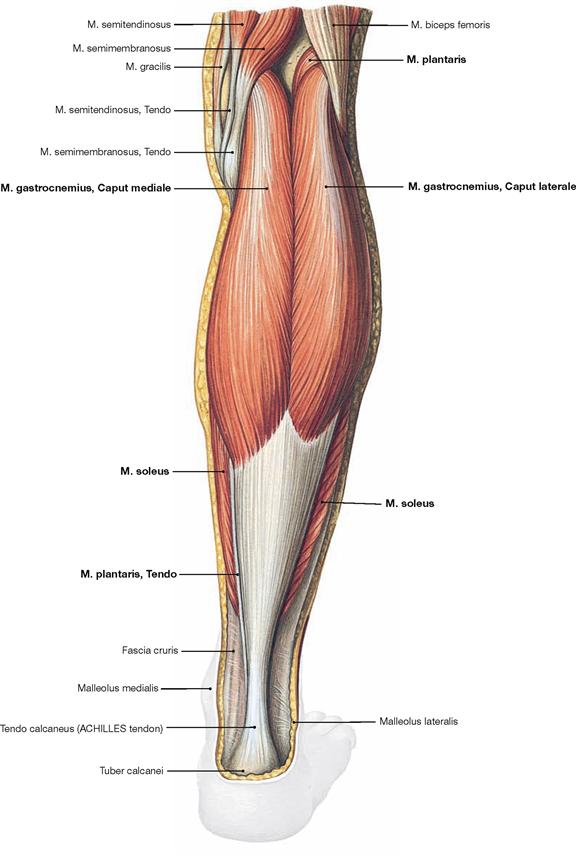
Fig. 4.136 Superficial layer of the dorsal muscles of the lower leg, right side; dorsal view.
The superficial group of flexors comprises the M. triceps surae and the M. plantaris. The strong M. triceps surae includes the two-headed M. gastrocnemius and the subjacent M. soleus. All superficial dorsal muscles insert at the Calcaneus via the ACHILLES tendon (Tendo calcaneus). The M. triceps surae is the strongest flexor of the ankle joint and the strongest supinator of the foot, even stronger than the M. tibialis posterior. If it is paralysed, such as after a disc herniation with resulting injury to the spinal cord segment S1 or a lesion of the N. tibialis, standing on one’s toes is impossible.
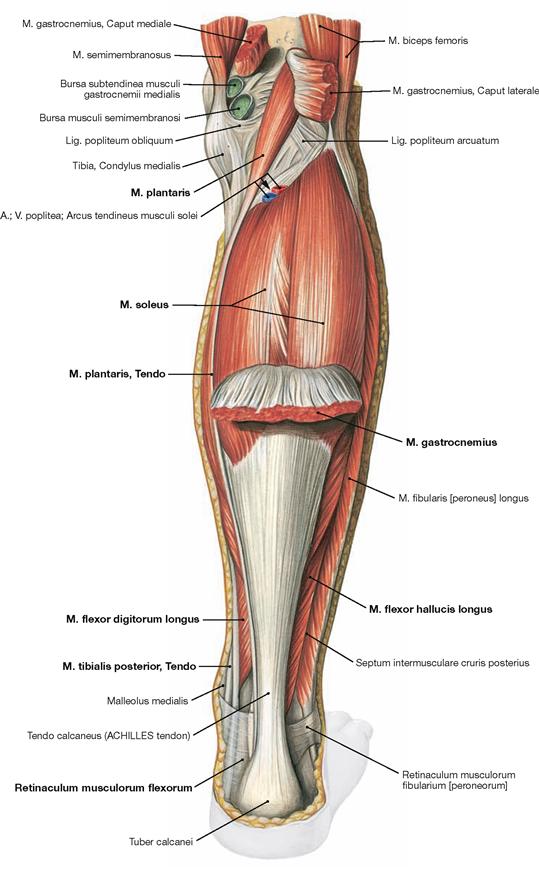
Fig. 4.137 Superficial layer of the dorsal muscles of the lower leg, right side; dorsal view; after dissecting the origins of the M. gastrocnemius.
After reflecting the M. gastrocnemius inferiorly, the M. plantaris is visible proximal of the M. soleus. The muscle bellies of the deep flexors are located further distally and are visible on both sides of the ACHILLES tendon after removal of the Fascia cruris. Their tendons are guided by the Retinaculum musculorum flexorum at the medial malleolus.
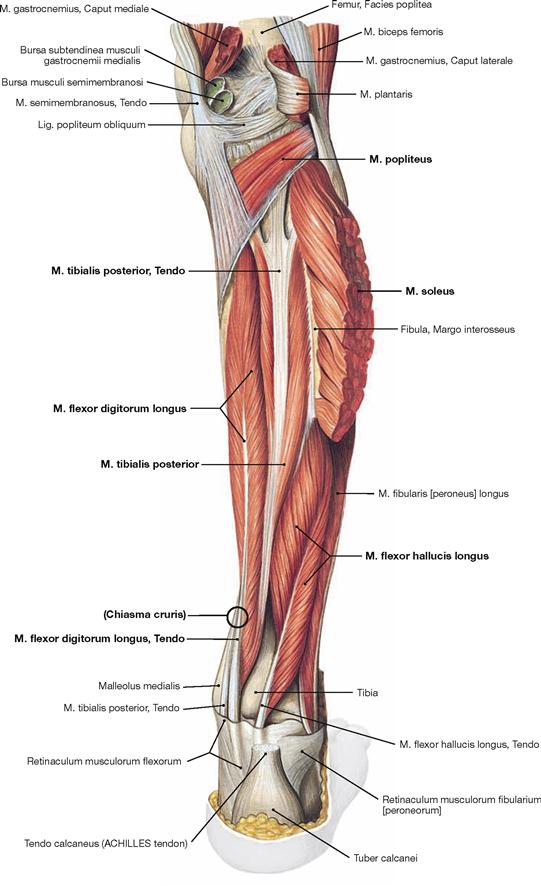
Fig. 4.138 Deep layer of the dorsal muscles of the leg, right side; dorsal view; after removal of the superficial flexors.
After removal of the superficial flexors the deep muscles are visible. The M. tibialis posterior is located between both flexor muscles of the toes. The M. flexor digitorum longus originates furthest medially followed by the M. tibialis posterior and further distal by the M. flexor hallucis longus. Their tendons meet beneath the medial malleolus where they are covered by the Retinaculum musculorum flexorum. In this course the tendon of the M. flexor digitorum longus crosses the tendon of the M. tibialis posterior (Chiasma cruris). Proximal, the M. popliteus originates from the Condylus lateralis and from the posterior horn of the lateral meniscus. The muscle inserts on the posterior aspect of the proximal Tibia and functions as a relatively strong medial rotator. Thus, the primary function of the M. popliteus is to actively stabilise the knee and to prevent an extensive lateral rotation.
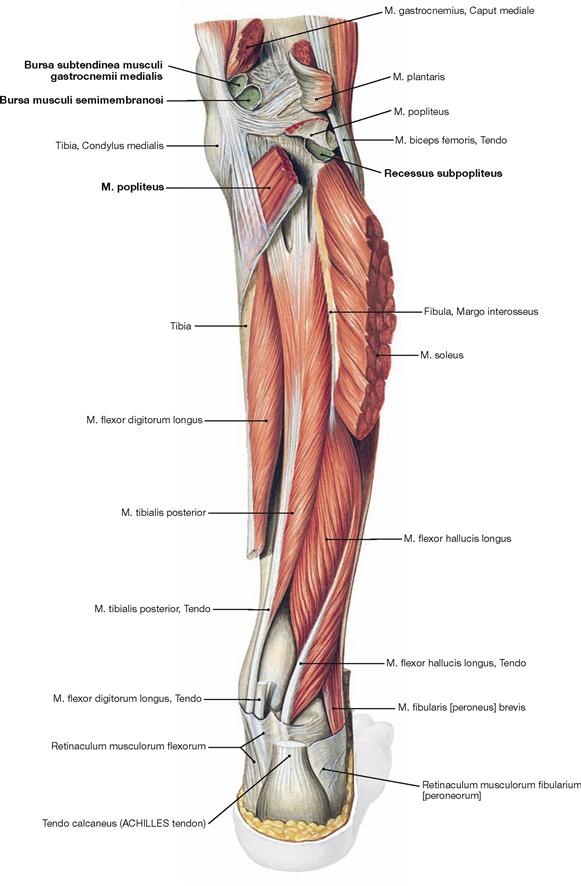
Fig. 4.139 Deep layer of the dorsal muscles of the leg, right side; dorsal view; after removal of the superficial flexors and splitting of the M. popliteus.
Upon splitting of the M. popliteus, the Bursa subpoplitea is exposed. This bursa frequently communicates with the joint cavity of the knee joint and is often referred to as Recessus subpopliteus. Additional bursae are present beneath the tendinous origins and insections of the dorsal muscles (Bursa musculi semimembranosi and Bursae subtendineae musculorum gastrocnemii medialis and lateralis). These also may communicate with the joint cavity (→ p. 280).
Synovial sheaths of the foot
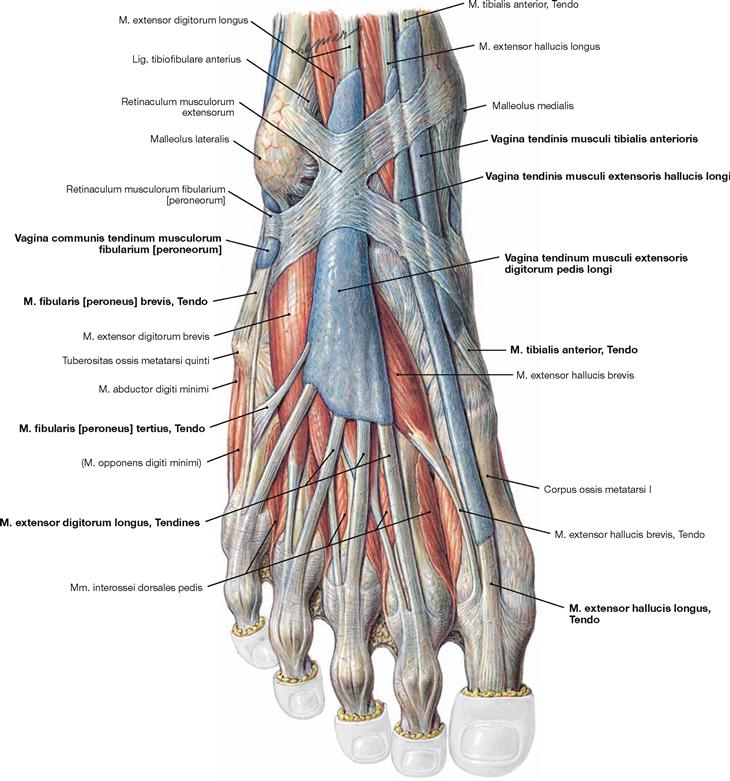
Fig. 4.140 Synovial sheaths, Vaginae tendinum, of the foot, right side; dorsal view in relation to the dorsum of the foot.
The Fascia cruris was removed except for the Retinaculum musculorum extensorum. The retinacula of the foot serve as retaining straps and prevent the tendons from lifting off the bones during muscle contractions. Each extensor muscle has its own synovial sheath (Vagina tendinis) which encloses all tendons of the respective muscle and serves as guiding tube as well as gliding surface. In contrast, the tendons of the M. fibularis longus and M. fibularis brevis have a common synovial sheath.
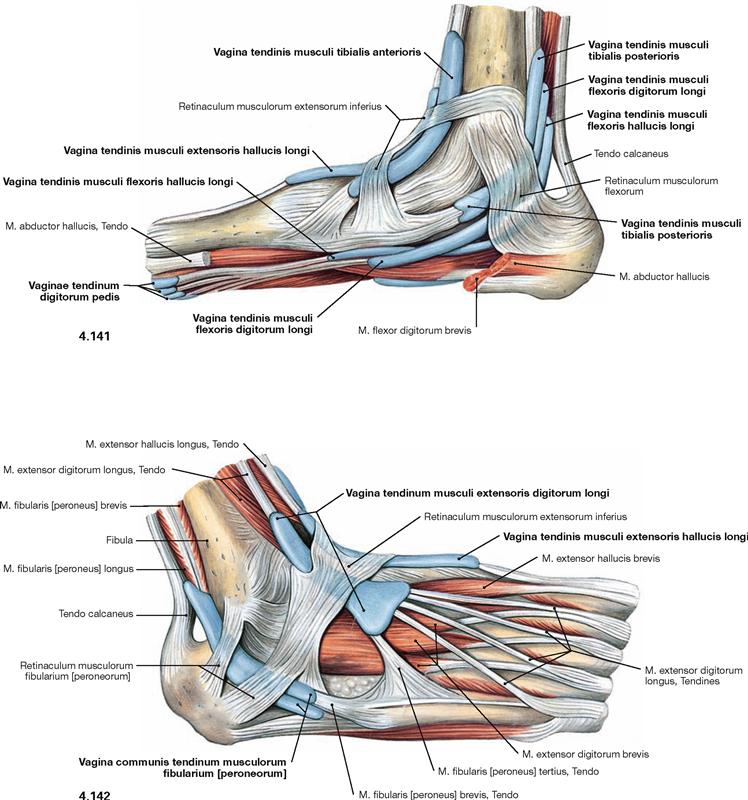
Fig. 4.141 and Fig. 4.142 Synovial sheaths, Vaginae tendinum, of the foot, right side; medial (→ Fig. 4.141) and lateral (→ Fig. 4.142) views.
The synovial sheaths surround the tendons of all three muscle groups of the leg particularly where the tendons are fixed to the bones by the retinacula. The Retinaculum musculorum flexorum forms the malleolar canal behind the medial malleolus which serves as a passageway for the neurovascular structures (N. tibialis, A./V. tibialis posterior) to the sole of the foot.
Muscles of the foot
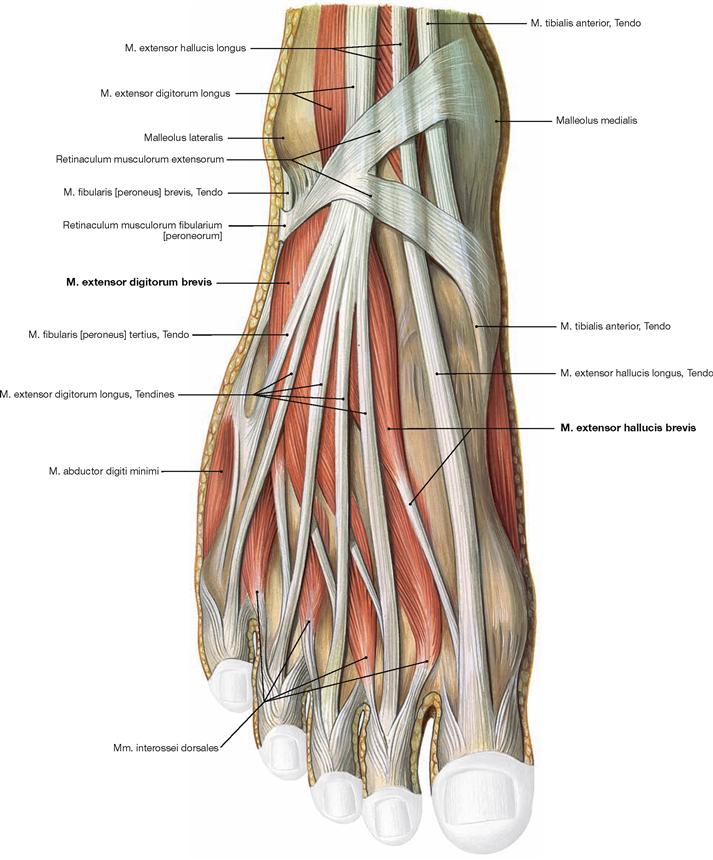
Fig. 4.143 Muscles of the dorsum of the foot, right side; dorsal view.
Beneath the tendons of the long extensor muscles, which have their muscle bellies at the ventral side of the leg, there are two short extensors. The M. extensor digitorum brevis and M. extensor hallucis brevis originate on the dorsal side of the Calcaneus and their tendons insert from lateral into the tendons of the long extensors and additionally into the dorsal aponeurosis. Therefore, they contribute to the extension in the phalangeal joints and the metatarsophalangeal joint of the big toe. The Mm. interossei dorsales are also visible, but they are grouped with the plantar muscles (→ p. 325).
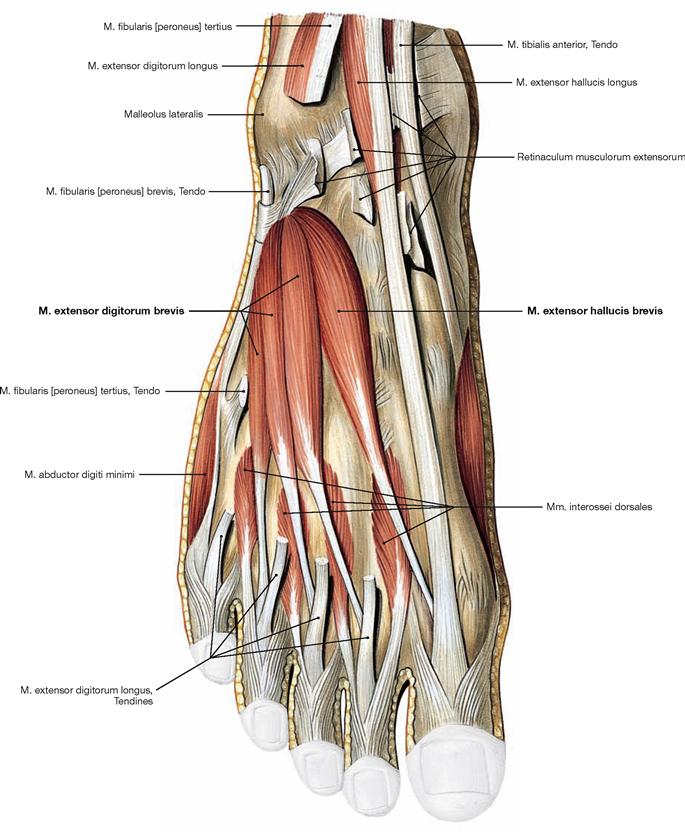
Fig. 4.144 Muscles of the dorsum of the foot, right side; dorsal view.
The Retinaculum musculorum extensorum was split and the tendon of the M. extensor digitorum longus partially removed to demonstrate the muscles of the dorsum of the foot. They comprise the short extensor muscles of the lateral four toes (M. extensor digitorum brevis) and of the big toe (M. extensor hallucis brevis). These muscles originate from the dorsal side of the Calcaneus and project to the dorsal aponeurosis of the second to fourth phalanges or to the dorsal side of the big toe.
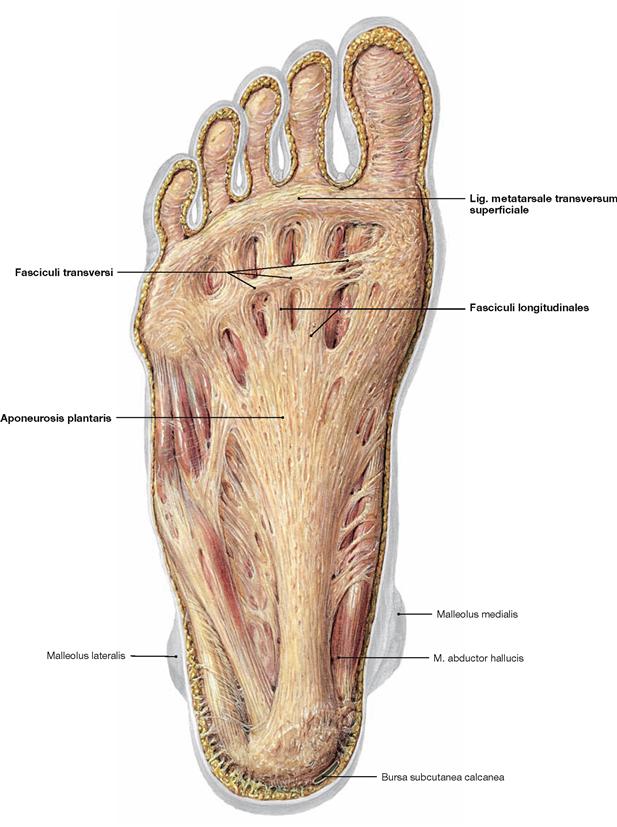
Fig. 4.145 Plantar aponeurosis, Aponeurosis plantaris, of the foot, right side; plantar view.
The plantar aponeurosis is a plate of dense connective tissue with a strong medial and two weaker lateral parts. The Fasciculi longitudinales project from the Tuber calcanei to the ligaments of the metatarsophalangeal joints. At the level of the Ossa metatarsi they are connected by transverse fibres (Fasciculi transversi). These transverse fibres are collectively referred to as Lig. metatarsale transversum superficiale. Two septa course from the plantar aponeurosis to the bones, thus, creating spaces for three plantar muscle groups.
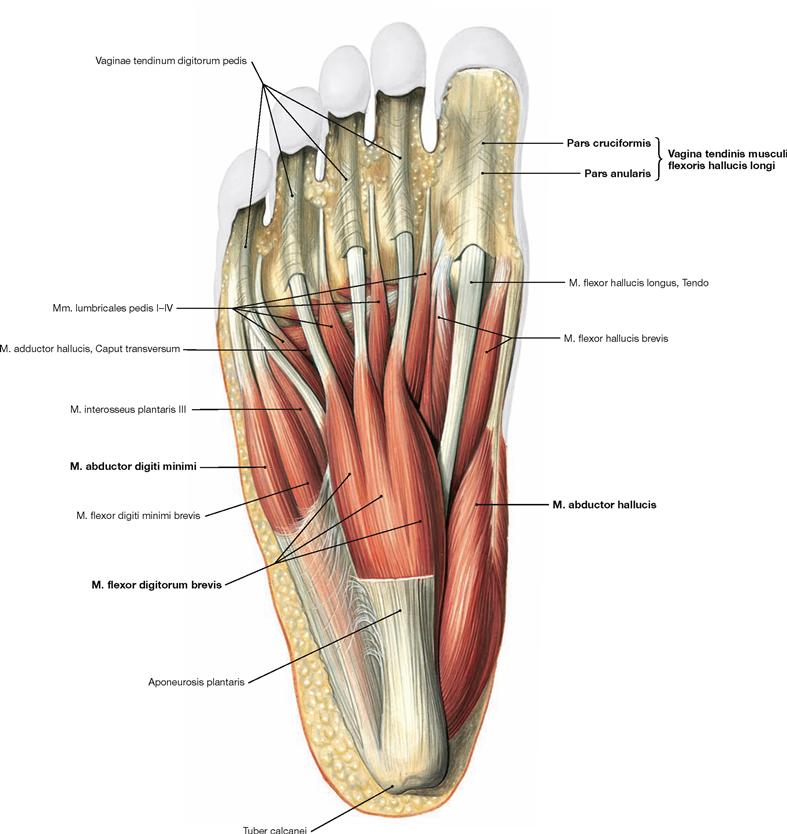
Fig. 4.146 Superficial layer of plantar muscles, right side; plantar view; after removal of the plantar aponeurosis.
In contrast to the hand, the muscles of the sole of the foot do not serve for differentiated movements of individual toes but serve in actively bracing the plantar arch as a functional muscle unit. The plantar muscles support the ligaments which accomplish a passive stabilisation. The plantar muscles are separated into three different groups (medial, intermediate and lateral) by two septa which project from the plantar aponeurosis to the bones. These groups are not easily separated during dissection, and it is easier to dissect four layers of muscles.
The muscles of the superficial layer comprise the M. abductor hallucis, M. flexor digitorum brevis and M. abductor digiti minimi. The tendons of the M. flexor digitorum brevis are pierced by the tendons of the long flexors. At the level of the toes, the tendons of the flexor muscles have separate synovial sheaths (Vaginae tendinum) which do not communicate with those at the tarsal level. The synovial sheaths are reinforced by ligaments containing anular (Pars anularis) and cruciform (Pars cruciformis) components.
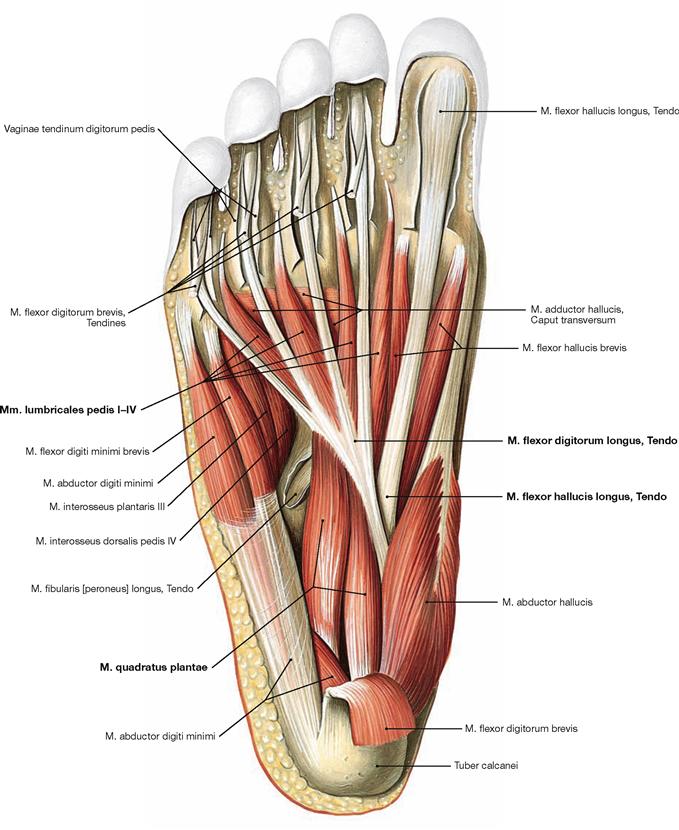
Fig. 4.147 Middle layer of the plantar muscles, right side; plantar view; after dissection of the M. flexor digitorum brevis.
The muscles lie in four layers on top of each other. Upon resection of the M. flexor digitorum brevis, the muscles and tendons of the second layer are visible. It consists of the tendons of the long flexors (M. flexor hallucis longus and M. flexor digitorum longus) and of two muscles of the intermediate group. The tendon of the M. flexor digitorum longus serves as origin for the M. quadratus plantae which functions as an accessory flexor supporting the long flexor muscle. The tendon also serves as origin for the four Mm. lumbricales which insert from medial on the proximal phalanges of the toes (II–V).
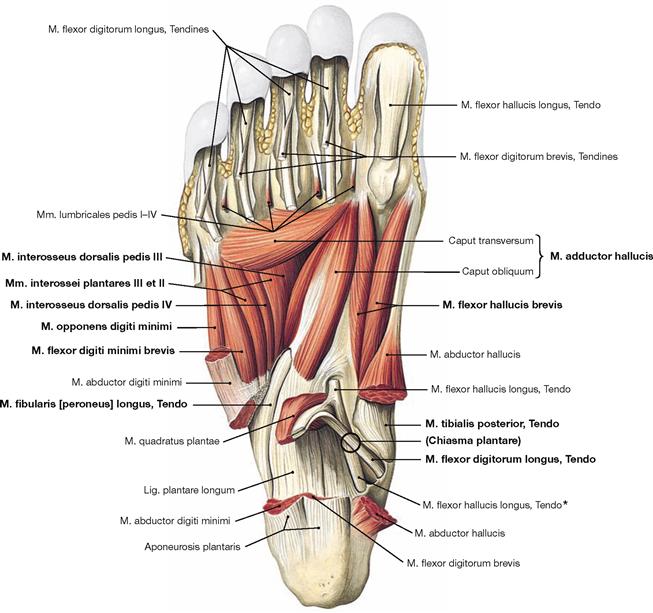
Fig. 4.148 Deep and deepest layers of the plantar muscles, right side; plantar view; after removal of both superficial muscle layers and the long flexor tendons.
Within the deep layer the M. flexor hallucis brevis and M. adductor hallucis are located medially, the M. flexor digiti minimi brevis and the inconsistent M. opponens digiti minimi laterally.
The deepest layer comprises three Mm. interossei plantares and four Mm. interossei dorsales as well as the tendons of the M. tibialis posterior and M. fibularis longus.
* The crossing of the M. flexor digitorum longus tendon over the M. flexor hallucis longus tendon is also referred to as Chiasma plantare.

Fig. 4.149 and Fig. 4.150 Mm. interossei dorsales (→ Fig. 4.149) and plantares (→ Fig. 4.150) of the foot, right side; dorsal (→ Fig. 4.149) and plantar (→ Fig. 4.150) views.
The four Mm. interossei dorsales (I–IV) are two-headed and originate from opposing sides of the bases of the Ossa metatarsi I to V. They insert on the proximal phalanges of the second to fourth toes in such a way that muscles I and II project medial and lateral to the second toe, whereas muscles III and IV course lateral to the third and fourth toe. Thus, the muscles can flex the metatarsophalangeal joints, abduct the toes II to IV laterally, and additionally adduct the second toe.
The three Mm. interossei plantares (I–III) have only one head and originate from the plantar side of the Ossa metatarsi III to V. They insert on the medial side of the respective toes. They serve for flexion of the metatarsophalangeal joints and adduction of the toes.
Plexus lumbosacralis
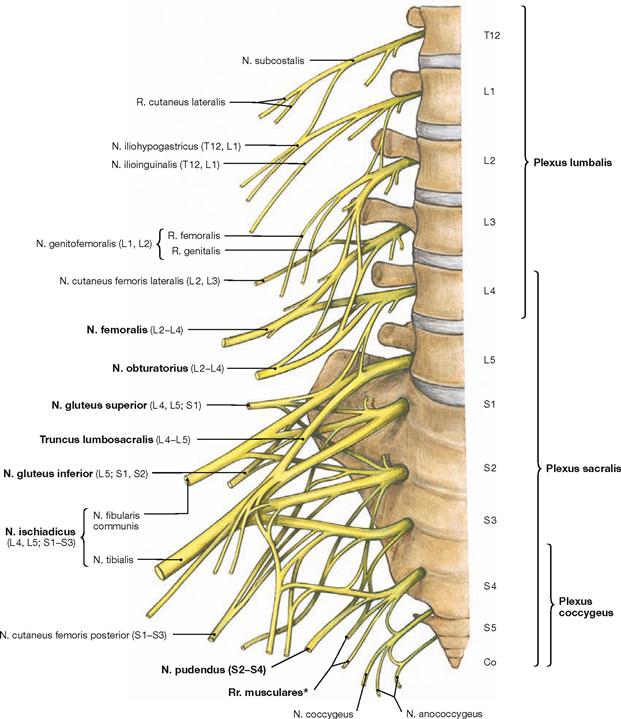
Fig. 4.151 Lumbosacral plexus, Plexus lumbosacralis (T12–S5, Co1): segmental organisation of the nerves, right side; ventral view.
The lower extremity is innervated by the Plexus lumbosacralis. The plexus is composed of Rr. anteriores of the spinal nerves which originate from the lumbar, sacral, and coccygeal segments of the spinal cord and combine to form the Plexus lumbalis (T12–L4) and the Plexus sacralis (L4–S5, Co1). The segments S4–Co1 are also referred to as Plexus coccygeus. Both plexuses are connected by the Truncus lumbosacralis which conveys nerve fibres from the spinal cord segments L4, L5 from the Plexus lumbalis to the small pelvis. The functionally most important nerves of the Plexus lumbalis are the N. femoralis and the N. obturatorius.
The N. femoralis provides motor innervation to the ventral muscle group of the hip and thigh (flexors in the hip and extensors in the knee) and sensory innervation to the ventral aspect of the thigh and the ventromedial aspect of the leg. The N. obturatorius conveys motor fibres to the adductor muscles and sensory fibres to the medial thigh. The strongest and longest branch of the Plexus sacralis is the N. ischiadicus. With both of its divisions (N. tibialis and N. fibularis) the N. ischiadicus provides motor innervation to the hamstring muscles (extensors in the hip and flexors in the knee) and to all muscles in the leg and the foot as well as sensory innervation to the calf and foot. The Nn. glutei superior and inferior innervate the gluteal muscles which represent the major extensors, rotators, and abductors of the hip. The N. pudendus provides motor innervation to the muscles of the perineal region and sensory innervation to the external genitalia. The muscles of the pelvic floor are innervated by direct branches (*) of the sacral plexus.
Topography
Innervation of the lower extremity by the Plexus lumbosacralis
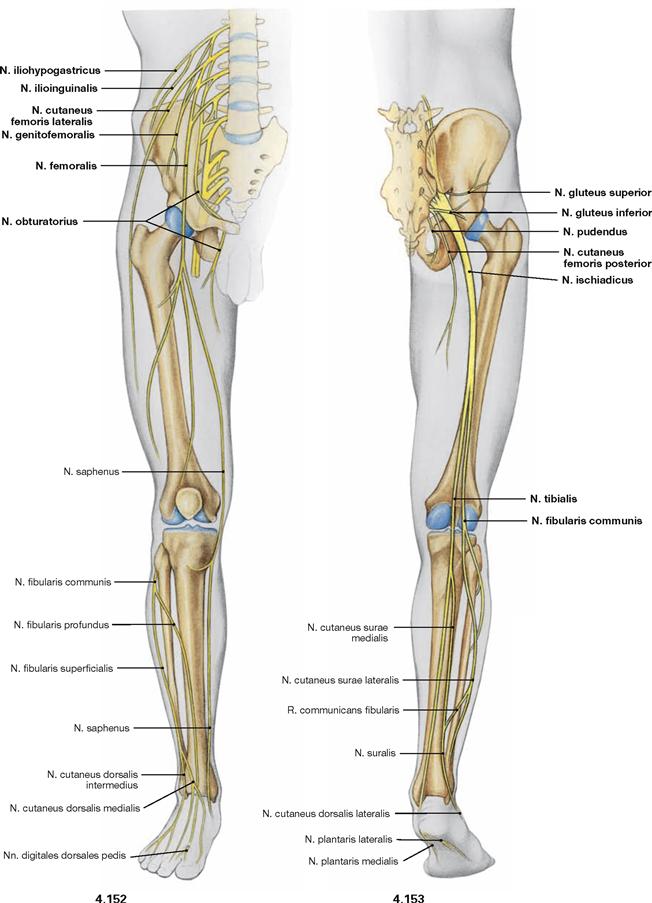
Fig. 4.152 and Fig. 4.153 Lumbosacral plexus, Plexus lumbosacralis (T12–S5, Co1): nerves of the lower extremity, right side; ventral (→ Fig. 4.152) and dorsal (→ Fig. 4.153) views.
The nerves of the Plexus lumbalis (T12–L4) course ventral to the hip joint and innervate the inferior part of the anterolateral abdominal wall and the ventral aspect of the thigh. The branches of the Plexus sacralis course dorsal to the hip joint. They innervate the posterior side of the thigh, the major part of the leg and the whole foot.
Innervation of the skin
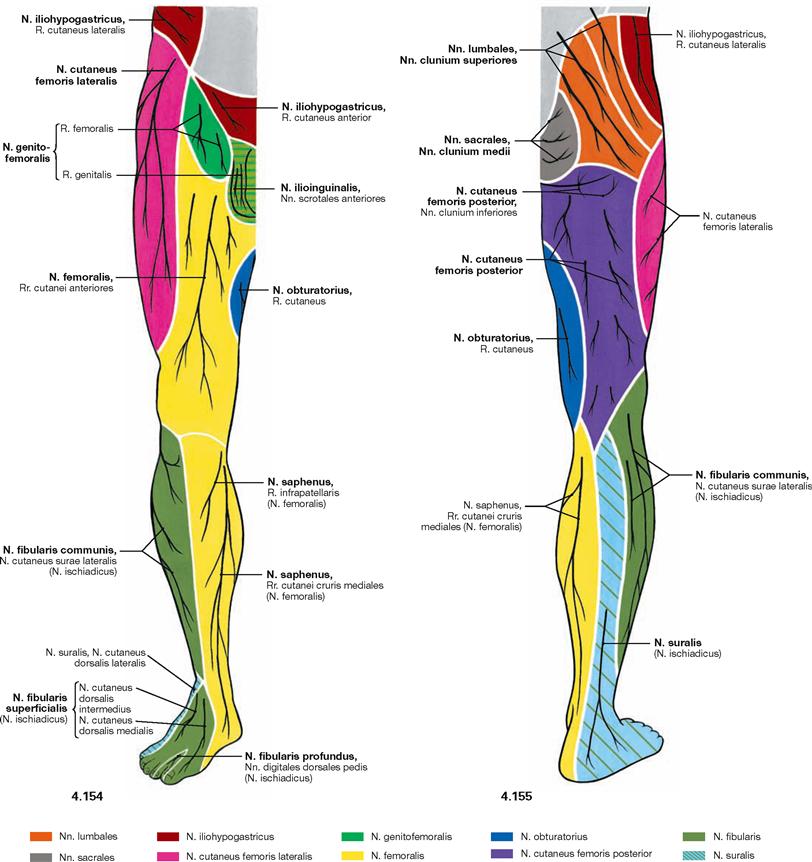
Fig. 4.154 and Fig. 4.155 Cutaneous nerves of the lower extremity, right side; ventral (→ Fig. 4.154) and dorsal (→ Fig. 4.155) views.
All nerves of the Plexus lumbalis contribute to the sensory innervation of the inguinal region and the ventral thigh. The lateral aspect of the leg and the dorsum of the foot are supplied by branches of the Plexus sacralis. The gluteal region is innervated by Rr. posteriores from the lumbar (Nn. clunium superiores) and sacral (Nn. clunium medii) spinal nerves. The dorsal side of the whole lower extremity and the sole of the foot are innervated by branches of the Plexus sacralis.
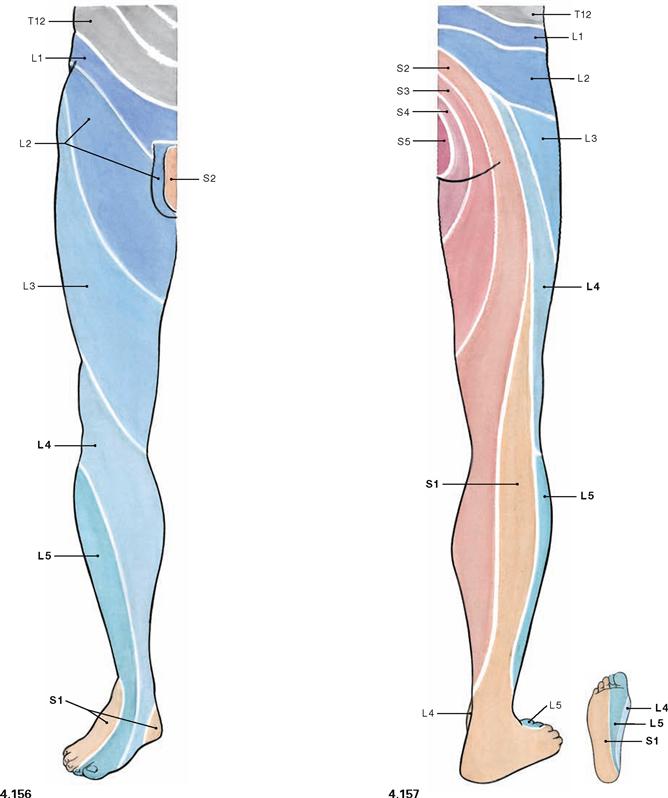
Fig. 4.156 and Fig. 4.157 Segmental innervation of the skin (dermatomes) of the lower extremity, right side; ventral (→ Fig. 4.156) and dorsal (→ Fig. 4.157) views.
Distinct areas of the skin are supplied by a single spinal cord segment. These cutaneous areas are referred to as dermatomes. Since the peripheral cutaneous nerves of the lower extremity convey sensory fibres from several spinal cord segments, the borders of the dermatomes do not correspond with the cutaneous area supplied by the peripheral nerves (→ p. 328). In contrast to the circular orientation of the dermatomes of the trunk, dermatomes on the ventral side of the lower extremity are obliquely oriented in a lateral superior to medial inferior direction. On the dorsal side they are oriented in a nearly longitudinal direction. (see Development, → p. 133).
Plexus lumbalis
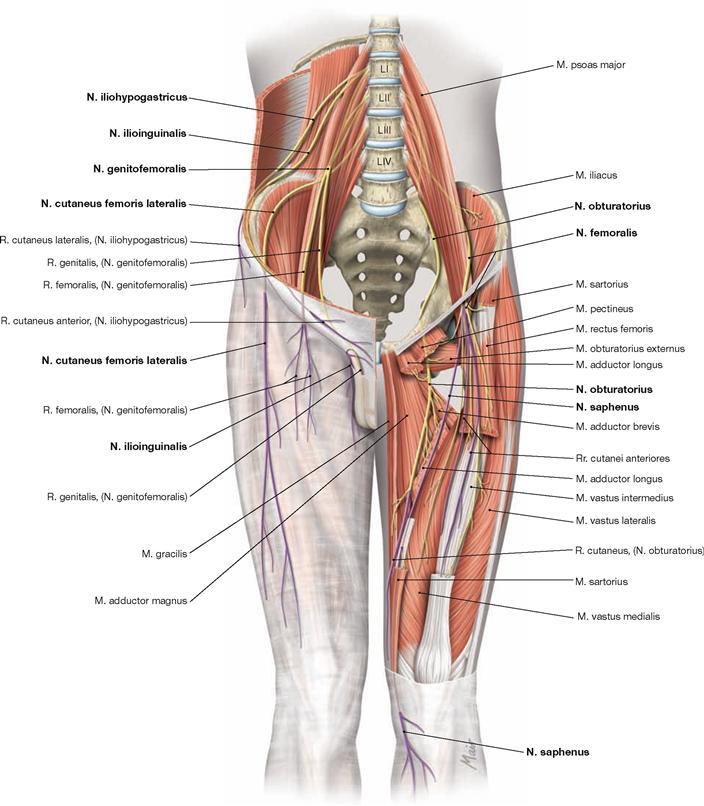
Fig. 4.158 Course and target areas of the nerves of the Plexus lumbalis (T12–L4); ventral view; cutaneous branches are highlighted in purple.
The N. iliohypogastricus and N. ilioinguinalis (further caudal) cross the M. quadratus lumborum behind the kidney and then pass between the M. transversus abdominis and the M. obliquus internus abdominis to the ventral side. Both innervate the inferior parts of these abdominal muscles. The N. iliohypogastricus also provides sensory innervation to the skin above the inguinal ligament, the N. ilioinguinalis provides sensory innervation to the anterior aspect of the external genitalia. The N. genitofemoralis pierces the M. psoas major, crosses posterior to the ureter, and divides into two branches: The lateral R. femoralis enters the anterior thigh through the Lacuna vasorum and provides cutaneous innervation inferior to the inguinal ligament. The medial R. genitalis courses through the inguinal canal to the Scrotum and conveys sensory fibres to the anterior aspects of the external genitalia and motor fibres to the M. cremaster in men. The N. cutaneus femoris lateralis projects laterally through the Lacuna musculorum and provides sensory fibres for the lateral side of the thigh. The N. femoralis courses medially through the Lacuna musculorum and immediately splits fan-like into several branches. Rr. cutanei anteriores supply the skin on the ventral side of the thigh. The Rr. musculares provide motor fibres to the anterior muscles of the hip (M. iliopsoas) and the thigh (M. sartorius and M. quadriceps femoris) and in part to the M. pectineus. Its terminal branch is the N. saphenus which enters the adductor canal (→ p. 351) and exits it through the Septum intermusculare vastoadductorium at the medial side of the knee joint to supply sensory innervation to the medial and anterior aspects of the leg. The N. obturatorius initially courses medial to the M. psoas major and then passes through the Canalis obturatorius (→ p. 351) to the medial aspect of the thigh. One of its branches reaches the M. obturatorius externus. The N. obturatorius then divides into the R. anterior and the R. posterior (anterior and posterior to the M. adductor brevis) which convey motor fibres to the muscles of the adductor group. The R. anterior also provides cutaneous innervation to the skin of the medial thigh.
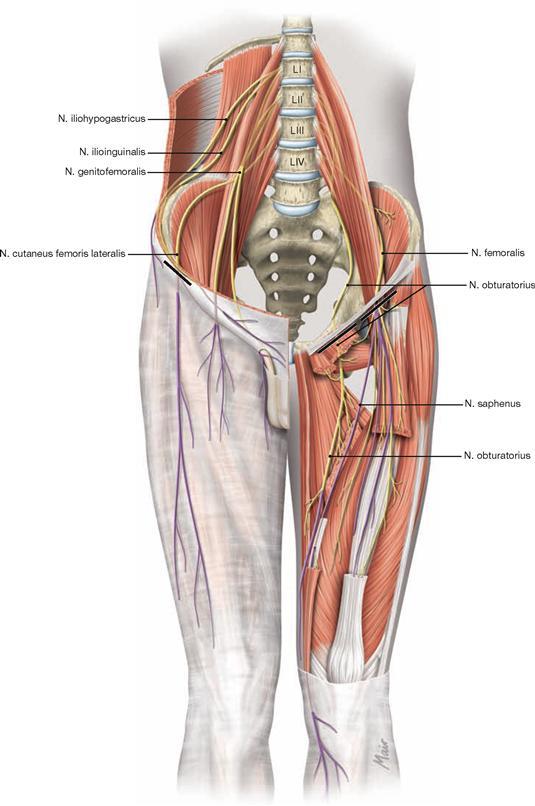
Fig. 4.159 Lesions of nerves of the Plexus lumbalis; ventral view. Cutaneous branches are highlighted in purple. Frequent locations for lesions are marked by black bars.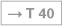
Plexus sacralis
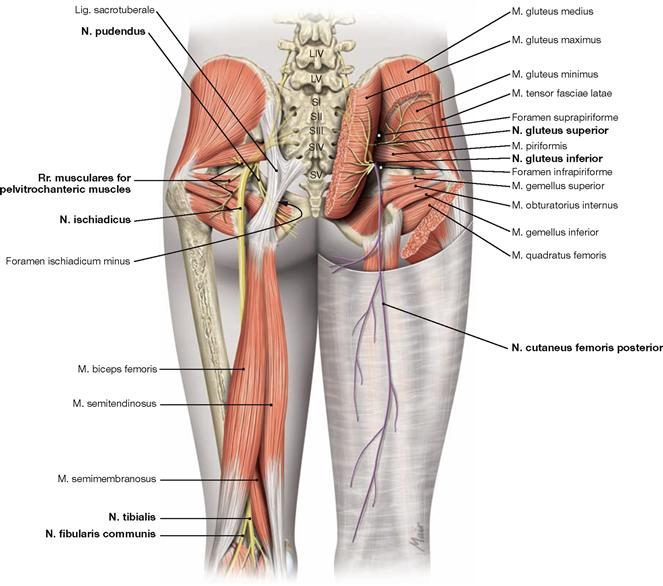
Fig. 4.160 Course and target areas of the nerves of the Plexus sacralis (L4–S5, Co1). Dorsal view; cutaneous nerves are highlighted in purple.
The N. gluteus superior exits the small pelvis through the Foramen suprapiriforme and provides motor innervation to the small gluteal muscles (most important abductors and medial rotators of the hip joint) and the M. tensor fasciae latae. The N. gluteus inferior exits through the Foramen infrapiriforme and innervates the M. gluteus maximus, the strongest extensor and external rotator of the hip joint.
The N. ischiadicus is the strongest nerve of the human body. It consists of two divisions (N. tibialis and N. fibularis communis) which are combined to one common nerve for a variable distance only by a connective tissue sheath (epineurium). The N. ischiadicus exits the pelvis through the Foramen infrapiriforme and descends to the popliteal fossa underneath the M. biceps femoris.
In most cases, N. tibialis and N. fibularis communis separate at the level of the distal third of the thigh. Occasionally (12% of cases), both nerves already exit the pelvis separately (high division) in which case the N. fibularis often pierces the M. piriformis. At the level of the thigh, the N. tibialis provides motor innervation to the hamstring muscles and the posterior head of the M. adductor magnus. The N. fibularis innervates the Caput breve of the M. biceps femoris. Both portions of the N. ischiadicus together innervate all muscles of the leg and the foot and provide sensory innervation to the skin of the leg (except for the medial aspect: innervated by the N. saphenus of the N. femoralis) and the foot.
The N. cutaneus femoris posterior exits the pelvis through the Foramen infrapiriforme and branches off the sensory Nn. clunium inferiores for the skin of the inferior gluteal region. It descends in the subfascial layer to the middle of the thigh and provides sensory innervation to the posterior thigh.
The N. pudendus has a complicated course. It exits the pelvis through the Foramen infrapiriforme and, together with the corresponding blood vessels, winds around the Spina ischiadica and courses through the Foramen ischiadicum minus medially into the ischioanal fossa. The N. pudendus courses in a fascial duplication of the M. obturatorius internus (ALCOCK’s canal; pudendal canal). The N. pudendus innervates the external sphincter muscle of the anal canal (M. sphincter ani externus) and all muscles of the perineum. It supplies sensory innervation to the posterior aspects of the external genitalia (posterior scrotum/labia majora; all of penis/clitoris).
The motor branches for the pelvitrochanteric muscles also exit through the Foramen infrapiriforme, whereas those for the pelvic floor do not exit the small pelvis. Parasympathetic nerves also remain within the pelvis. The small cutaneous branches pierce the Lig. sacrotuberale (N. cutaneus perforans) or the M. ischiococcygeus (N. anococcygeus).
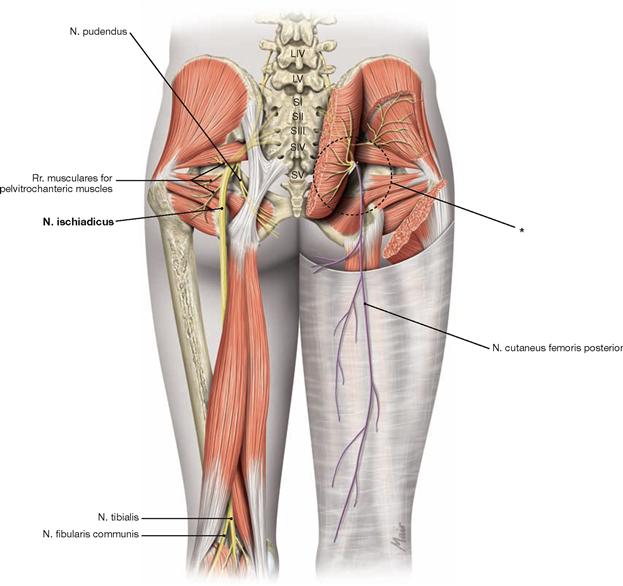
Fig. 4.161 Lesions of the most important nerves of the Plexus sacralis. Dorsal view. Cutaneous branches are highlighted in purple.
On the right side, the potential injury, such as wrongly placed intragluteal injections, to the nerves at the level of their exit from the pelvis is shown. On the left side, the possible site of injury to the N. ischiadicus due to fractures of the pelvis or hip surgery is indicated.
* lesion with wrongly placed intragluteal injection
Intragluteal injections
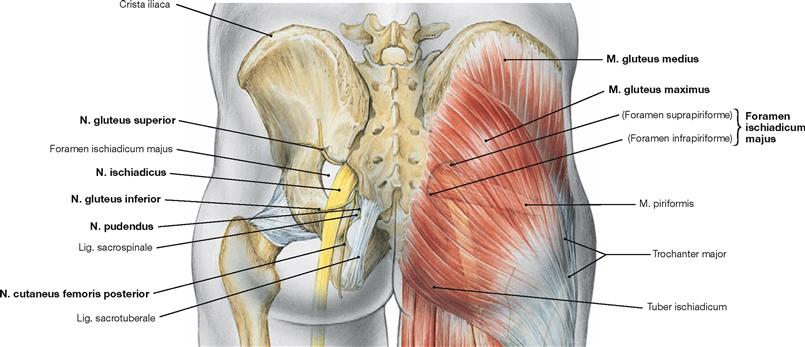
Fig. 4.162 Surface projection of the skeletal contour and the N. ischiadicus in the gluteal region.
With wrongly positioned intragluteal injections in the M. gluteus maximus principally all neurovascular structures passing through the Foramen ischiadicum majus are at risk of injury. Only the A. and V. pudenda interna and the N. pudendus are well protected as then course medially and pass through the Foramen ischiadicum minus to reach the ischioanal fossa. Therefore, injections should always be applied into the M. gluteus medius (→ Fig. 4.163).
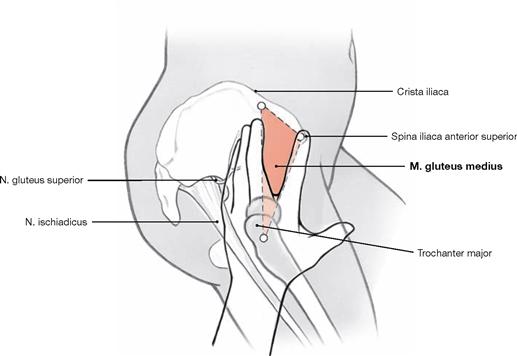
Fig. 4.163 Ventral intragluteal injections (according to HOCHSTETTER)
To avoid damaging of important neurovascular structures in the gluteal region, intragluteal injections are performed within a triangular field between two splayed fingers and the Crista iliaca. The index finger is placed onto at the Spina iliaca anterior superior and the palm of the hand over the Trochanter major. The only nerve remaining at risk is the motor branch projecting from the N. gluteus superior to the M. tensor fasciae latae.
Plexus sacralis
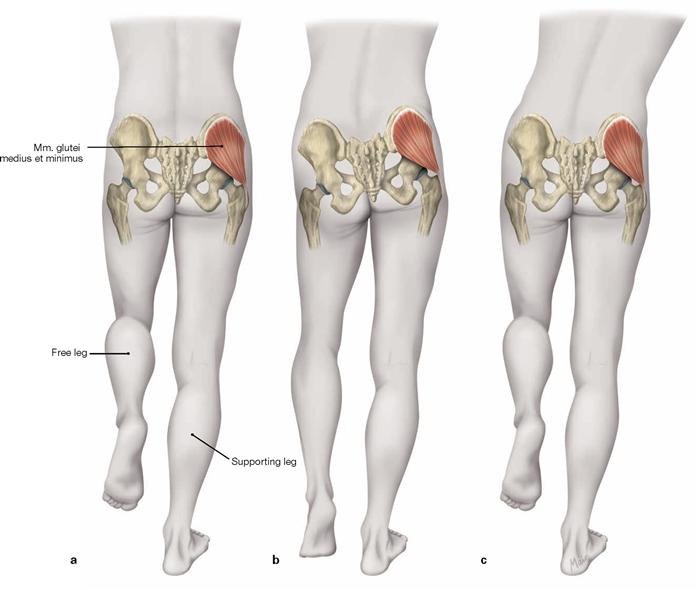
Figs. 4.164a to c TRENDELENBURG’s sign and DUCHENNE’s gait with loss of function of the small gluteal muscles on the right side (b, c).
a. The gluteal muscles abduct the ipsilateral leg if the body weight is shifted to the other leg. In one-legged stand, the ipsilateral muscles stabilise the pelvis and prevent the tilting of the pelvis to the contralateral side.
b. With functional insufficiency of the small gluteal muscles, such as in hip dysplasia or with lesions of the N. gluteus superior, the pelvis drops to the healthy side when standing on the leg of the affected side (TRENDELENBURG’s sign).
c. The pelvis of the healthy side is elevated by shifting the trunk towards the affected side (DUCHENNE’s gait).
N. tibialis
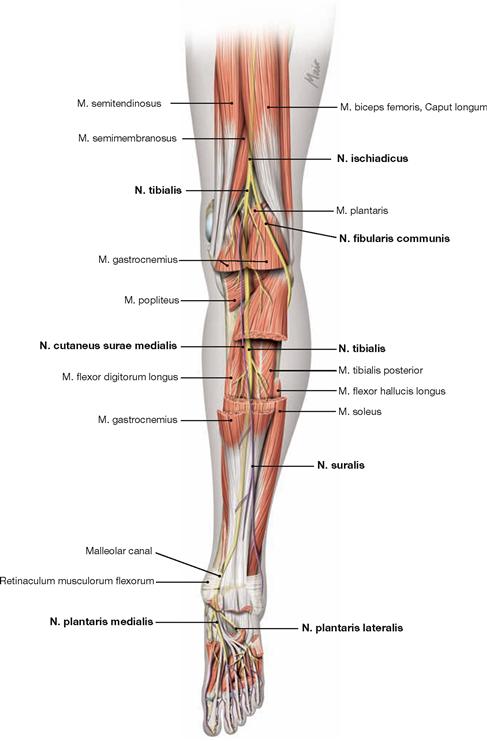
Fig. 4.165 N. tibialis: sensory innervation by cutaneous nerves (purple), and motor innervation by muscular branches, right side; dorsal view.
The N. ischiadicus often divides at the transition from the middle to the lower third of the thigh into the medial N. tibialis and the lateral N. fibularis communis. The N. tibialis innervates the dorsal muscles of the thigh (hamstring muscles and dorsal part of the M. adductor magnus). The N. tibialis continues in the direction of the N. ischiadicus to pass the popliteal fossa and descends between the heads of the M. gastrocnemius beneath the tendinous arch of the M. soleus (Arcus tendineus musculi solei). It further courses together with the A. and V. tibialis posterior between the superficial and deep flexors to the medial malleolus. In the popliteal fossa the N. cutaneus surae medialis branches off to supply the medial calf and splits into the N. suralis for the distal calf and the N. cutaneus dorsalis lateralis for the lateral margin of the foot. The latter often communicates with a cutaneous branch from the N. fibularis communis. When passing underneath the Retinaculum musculorum flexorum (malleolar canal), the N. tibialis splits into its two terminal branches (Nn. plantares medialis and lateralis) for the innervation of the sole of the foot. Thus, the N. tibialis provides motor innervation to all flexor muscles of the calf and all plantar muscles of the foot as well as sensory innervation to the middle calf and, after forming the N. suralis, to the lower calf and the lateral margin of the foot.
N. fibularis communis
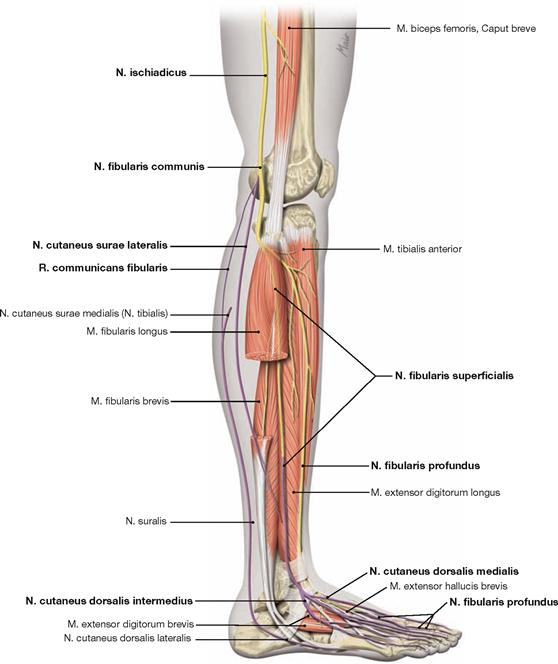
Fig. 4.166 N. fibularis communis: sensory innervation by cutaneous nerves (purple), and motor innervation by muscular branches, right side; lateral view.
After the division of the N. ischiadicus at the transition to the distal third of the thigh, the N. fibularis communis courses through the popliteal fossa and around the head of the fibula to the fibularis compartment. Here the nerve divides into its two terminal branches (Nn. fibulares superficialis and profundus). At the thigh, the N. fibularis communis provides motor innervation to the Caput breve of the M. biceps femoris only. Prior to its division into the terminal branches, the N. fibularis communis provides the N. cutaneus surae lateralis for the skin of the lateral calf and another branch for the communication with the N. cutaneus surae medialis.
The N. fibularis superficialis continues in the fibularis compartment and provides motor fibres to the fibularis muscles. Subsequently, it pierces the fascia of the distal leg and splits into the two terminal sensory branches (Nn. cutanei dorsales medialis and intermedius) for the dorsum of the foot.
The N. fibularis profundus enters the extensor compartment and descends together with the A. tibialis anterior to the dorsum of the foot. On its way, it provides motor innervation to the extensor muscles of the leg and the dorsum of the foot. Its terminal branch and provides sensory innervation to the skin of the first interphalangeal space.
Arteries of the pelvis and thigh
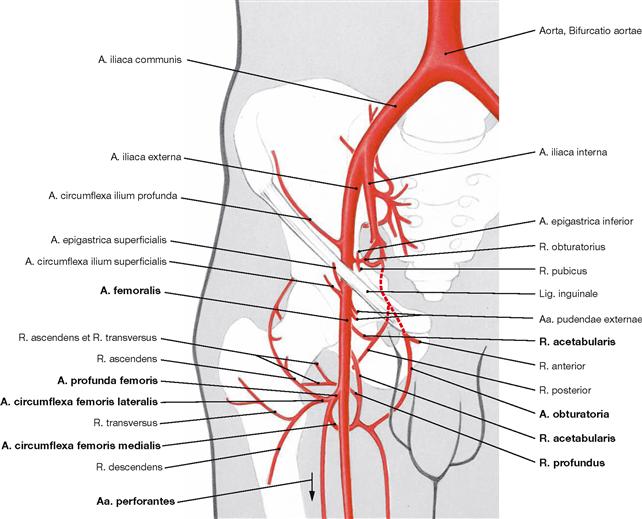
Fig. 4.167 Arteries of the pelvis and the thigh, right side; ventral view.
The A. profunda femoris is the main artery of the hip joint and the thigh. The other branches of the A. femoralis do not contribute to the arterial supply of the thigh. The A. profunda femoris branches off the A. femoralis 3–6 cm inferior to the inguinal ligament and divides into the Aa. circumflexae femoris medialis and lateralis. In the adult, the femoral head is almost exclusively supplied by the A. circumflexa femoris medialis (R. profundus) which loops around the Collum femoris from behind (→ Figs. 4.57 und 4.58). The R. profundus also supplies the adductor muscles and the hamstring muscles. The R. acetabularis anastomoses with the identically named branch of the A. obturatoria. The A. circumflexa femoris lateralis courses anterior to the femoral neck. It supplies the femoral neck and with several branches also the lateral hip muscles and the ventral muscles of the thigh. The Aa. perforantes are terminal branches which supply the adductor and hamstring muscles. All branches anastomose with each other as well as with the A. obturatoria and the Aa. gluteae from the A. iliaca interna which is the basis for potential collateral circulations.
Arteries of the lower extremity
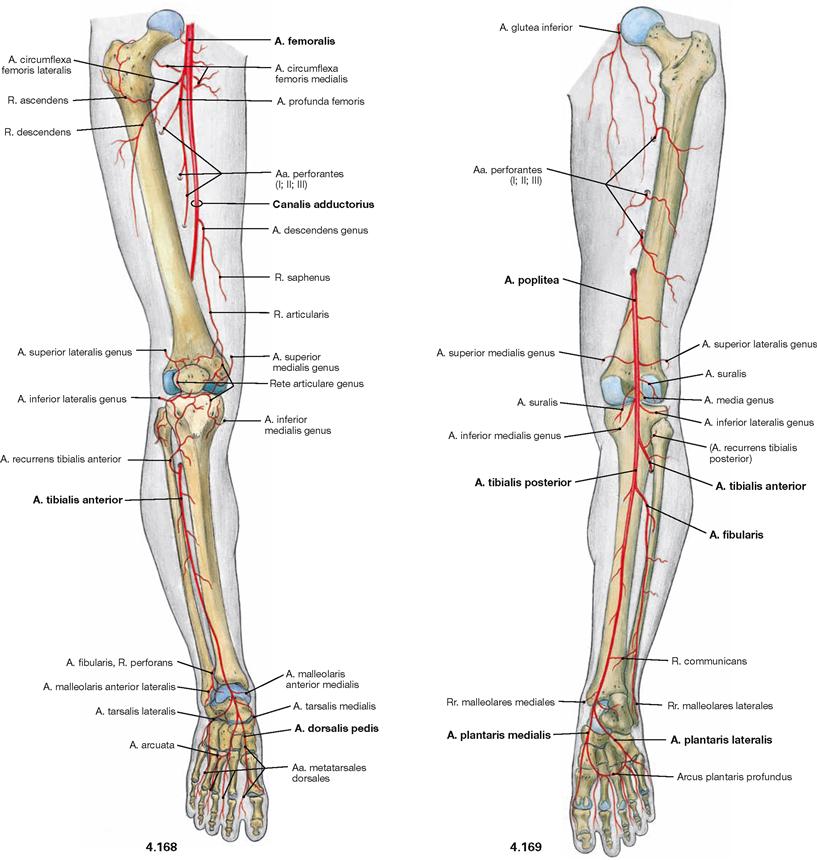
Fig. 4.168 and Fig. 4.169 Arteries of the lower extremity, right side; ventral (→ Fig. 4.168) and dorsal (→ Fig. 4.169) views.
The A. iliaca externa branches off the A. iliaca communis anterior to the sacro-iliac joint and continues beneath the inguinal ligament in the Lacuna vasorum as A. femoralis. Following the passage through the adductor canal it is then referred to as A. poplitea (arterial supply of the knee joint). The A. poplitea descends underneath the tendinous arch of the M. soleus between the superficial and deep flexors of the leg and divides into the A. tibialis posterior which continues its course, and the A. tibialis anterior which pierces the Membrana interossea cruris to reach the anterior extensor compartment. The latter continues as A. dorsalis pedis on the dorsum of the foot. The A. tibialis posterior provides the strong A. fibularis to the lateral malleolus and then continues through the malleolar canal around the medial malleolus to reach the sole of the foot, where it provides two terminal branches (Aa. plantares medialis and lateralis).
Veins of the lower extremity
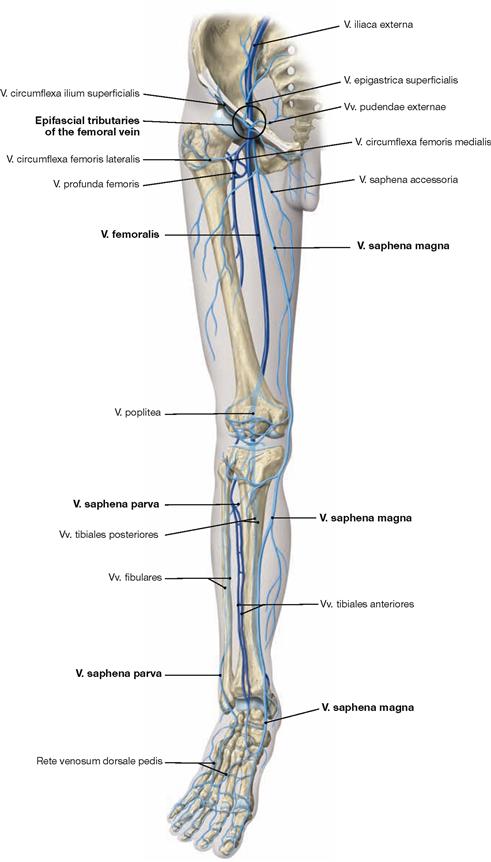
Fig. 4.170 Veins of the lower extremity, right side; ventral view.
The deep veins (dark blue) accompany the respective arteries. In the leg, usually two veins course together with the respective artery, whereas at the thigh and the popliteal fossa only one concomitant vein is found. The superficial venous system (light blue) consists of two main veins which collect the blood from the dorsum and the sole of the foot.
The V. saphena magna originates anterior to the medial malleolus and ascends on the medial side of the leg and thigh to the Hiatus saphenus (→ Fig. 4.178). Here, the V. saphena magna receives tributaries from several veins of the inguinal region (see below) and enters the V. femoralis at the femoral triangle.
On the posterior side, the V. saphena parva originates from the lateral margin of the foot posterior to the lateral malleolus and ascends on the middle of the calf to the popliteal fossa to enter the V. poplitea. The V. saphena magna and parva communicate through variable branches.
Tributaries of the V. saphena magna at the femoral triangle:
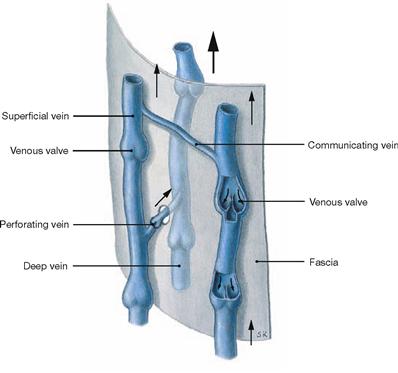
Fig. 4.171 Superficial and deep veins of the lower extremity with venous valves: organisation principle.
The extremities have a superficial epifascial venous system and a deep subfascial venous system coursing together with the respective arteries. Both systems are connected by perforating veins (Vv. perforantes). Venous valves direct the blood flow from the superficial towards the deep veins causing the major part of the blood (85%) to be drained via the deep veins of the lower extremity to the heart. Among the many perforating veins, three groups are of clinical relevance:
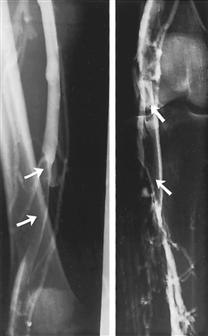
Lymph vessels of the lower extremity
Fig. 4.173 and Fig. 4.174 Superficial lymph vessels of the lower extremity, right side; ventral (→ Fig. 4.173) and dorsal (→ Fig. 4.174) views.
Alongside the veins there are a superficial and a deep system of collecting lymph vessels with incorporated lymph nodes. The superficial ventromedial system alongside the V. saphena magna is the main lymphatic drainage of the lower extremity and drains into the superficial inguinal nodes (Nodi lymphoidei inguinales superficiales) (→ p. 344). The smaller dorsolateral system parallels the V. saphena parva and drains into the lymph nodes of the popliteal fossa (Nodi lymphoidei poplitei superficiales and profundi) and continues into the deep inguinal lymph nodes (Nodi lymphoidei inguinales profundi). The deep collecting systems directly drain into the deep popliteal and inguinal lymph nodes.
While most of the venous drainage from the lower extremity occurs via the deep veins, the major part of the lymph is drained by the superficial lymph vessels.
Lymph nodes and lymph vessels of the inguinal region
Fig. 4.175 and Fig. 4.176 Superficial lymph nodes of the inguinal region, Regio inguinalis (→ Fig. 4.175), and their tributaries (→ Fig. 4.176), right side; ventral view.
The inguinal region harbours four to 25 epifascial superficial inguinal lymph nodes (Nodi lymphoidei inguinales superficiales) which further drain into one to three inguinal lymph nodes medial of the V. femoralis (Nodi lymphoidei inguinales profundi), and further into the Nodi lymphoidei iliaci externi in the pelvis. The superficial inguinal lymph nodes form a vertical strand along the V. saphena magna and a horizontal strand beneath the inguinal ligament.
The inguinal lymph nodes not only serve as regional lymph nodes for the major part of the lower extremity but also collect lymph from the lower quadrants of the abdominal wall and the back, the perineal region and the external genitalia (→ Figs. 2.111 to 2.114). In addition, lymph from the lower parts of the Rectum and the Vagina and occasionally from the Uterus and adjacent uterine tubes (along the Lig. teres uteri) drains into the inguinal lymph nodes.
Vessels and nerves of the inguinal region
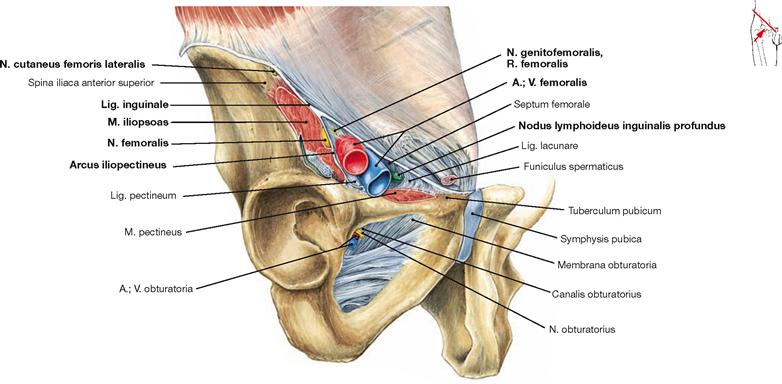
Fig. 4.177 Lacunae musculorum and vasorum, right side; oblique section at the level of the inguinal ligament; ventral view.
The space between Os coxae and the Lig. inguinale (Fossa iliopectinea) is divided by the Arcus iliopectineus, which spans between the inguinal ligament and the pelvic bone, into the lateral Lacuna musculorum and the medial Lacuna vasorum. The Lacuna musculorum is almost completely occupied by the M. iliopsoas. The N. cutaneous femoris lateralis is located lateral to the M. iliopsoas near the Spina iliaca anterior, the N. femoralis is positioned medial to the M. iliopsoas. Passing through the Lacuna vasorum from lateral to medial are the R. femoralis of the N. genitofemoralis, the A. femoralis, and the V. femoralis. Located most medially are the deep inguinal lymph nodes (Nodi lymphoidei inguinales profundi).
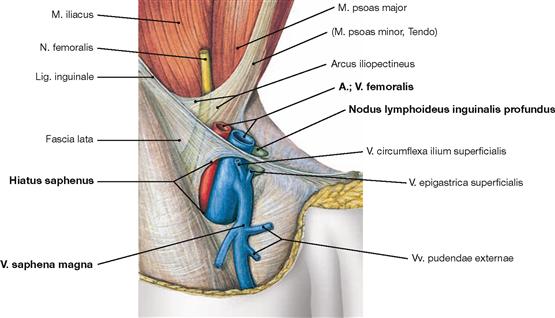
Fig. 4.178 Hiatus saphenus and Lacuna vasorum, right side; ventral view; after removal of the anterior abdominal wall, of the Fascia iliaca and the abdominal viscera.
The Hiatus saphenus is an opening of the Fascia lata through which the V. saphena magna passes prior to entering the V. femoralis. Located most medially are the deep inguinal lymph nodes (Nodi lymphoidei inguinales profundi) the biggest of which is referred to as ROSENMUELLER’s node.
Superficial vessels and nerves of inguinal region and thigh
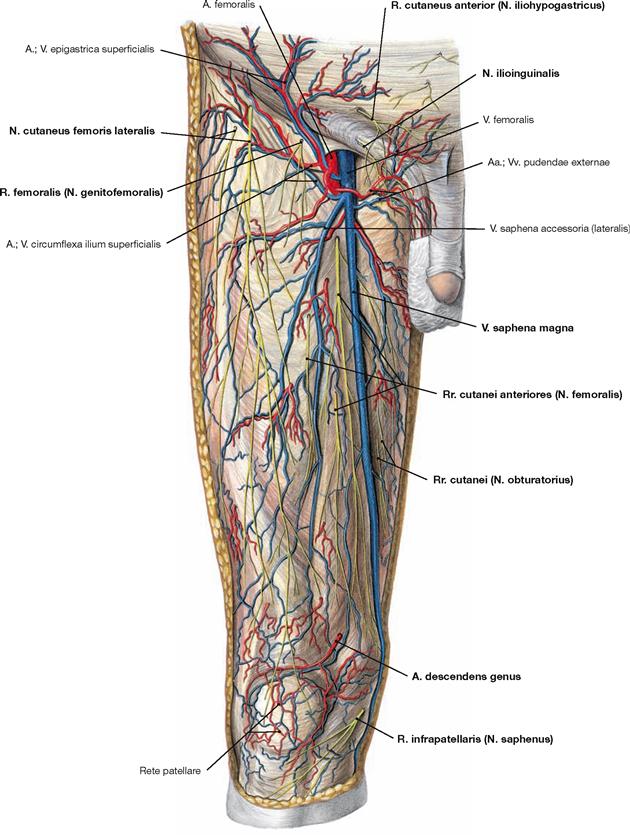
Fig. 4.179 Epifascial vessels and nerves of the inguinal region, Regio inguinalis, the thigh, Regio femoris anterior, and the knee, Regio genus anterior, right side; ventral view.
During the dissection of this region particularly the course of the cutaneous nerves and the epifascial veins need to be considered. The N. ilioinguinalis pierces the fascia above the inguinal ligament. Just cranial to it, the R. cutaneus anterior of the N. iliohypogastricus is found. The V. saphena magna ascends at the medial aspect of the thigh and enters into the V. femoralis through the Hiatus saphenus. Here the vein collects several tributaries from the inguinal region (→ p. 341). Most of these veins are accompanied by small branches of the A. femoralis. The R. femoralis of the N. genitofemoralis passes through the Lacuna vasorum just lateral to the A. femoralis. The N. cutaneus femoris lateralis traverses the Lacuna musculorum medially to the Spina iliaca anterior superior and innervates the lateral aspect of the thigh. The Rr. cutanei anteriores of the N. femoralis pierce the fascia at several locations to innervate the ventral aspect of the thigh. Medial to the V. saphena magna, several small cutaneous branches of the N. obturatorius supply a variable area on the medial aspect of the thigh. Medial and inferior to the knee, the R. infrapatellaris of the N. saphenus passes through the fascia. Just above the patella the thin A. descendens genus courses to the Rete patellare of the knee.
Superficial vessels and nerves of the gluteal region and thigh
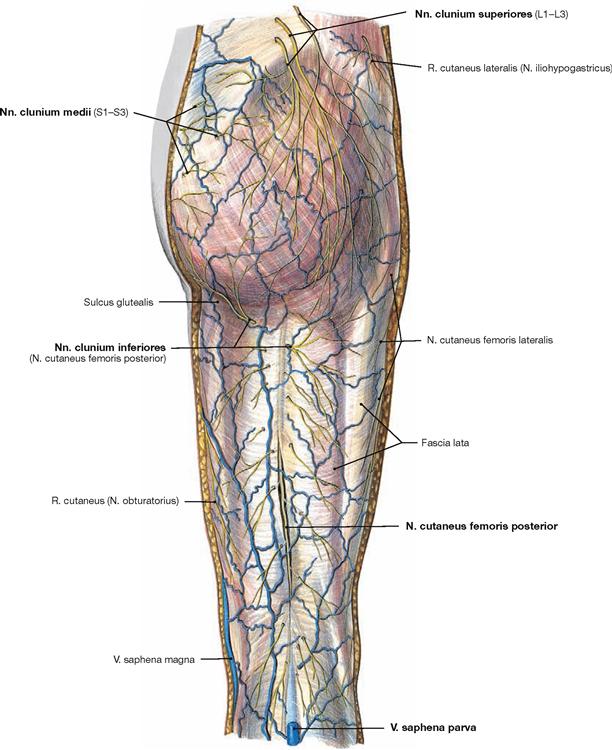
Fig. 4.180 Epifascial vessels and nerves of the gluteal region, Regio glutealis, the thigh, Regio femoris posterior, and the popliteal fossa, Fossa poplitea, right side; dorsal view.
There are no important epifascial veins on the posterior side of the thigh. The V. saphena parva of the leg enters the subfascial V. poplitea in the popliteal fossa. The skin of the gluteal region is innervated by three groups of cutaneous nerves. The Nn. clunium superiores (Rr. posteriores from L1–L3) appear laterally to the autochthonous muscles of the back above the iliac crest. The Nn. clunium medii (Rr. posteriores from S1–S3) traverse the M. gluteus maximus at its origin from the posterior side of the sacrum. The Nn. clunium inferiores are branches of the N. cutaneus femoris posterior and wind around the inferior margin of the M. gluteus maximus. The N. cutaneus femoris posterior descends in the middle of the thigh and pierces the fascia at midlength of the thigh to provide sensory innervation to the posterior aspect of the thigh.
Superficial vessels and nerves of the leg
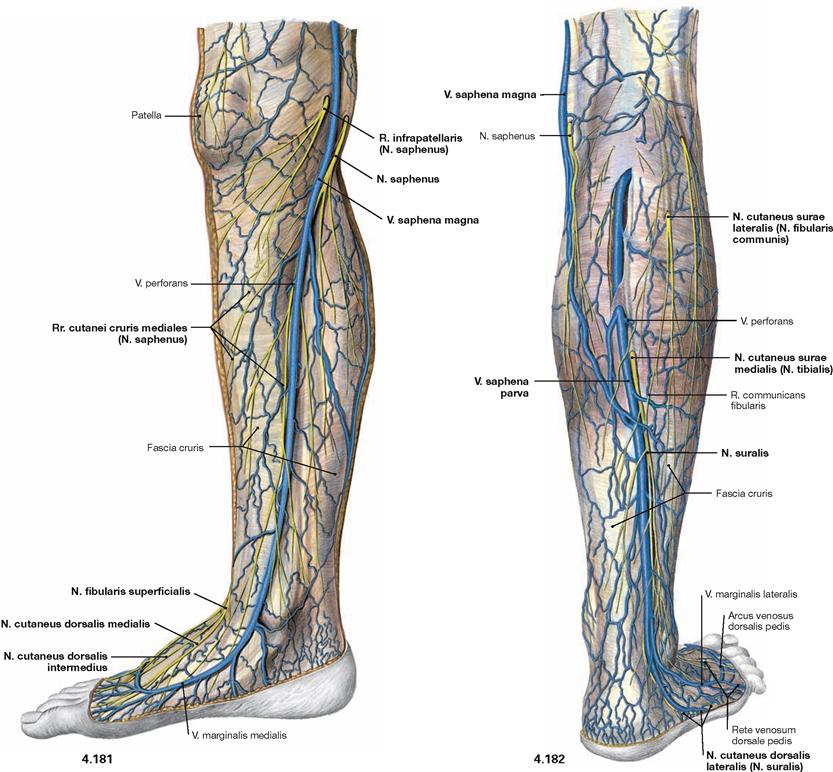
Fig. 4.181 and Fig. 4.182 Epifascial veins and nerves of the leg, Regio cruris, and the foot, Regio pedis, right side; medial (→ Fig. 4.181) and dorsolateral (→ Fig. 4.182) views.
The V. saphena magna originates at the medial margin of the foot anterior to the medial malleolus and ascends on the medial side of the leg and thigh. At the medial aspect of the knee, the N. saphenus pierces the fascia. Its major branch descends adjacent to the V. saphena magna and splits into the sensory terminal branches, Rr. cutanei cruris mediales, for the innervation of the ventral and medial leg and the medial margin of the foot. The R. infrapatellaris of the N. saphenus pierces the fascia ventral to the V. saphena magna and supplies the skin beneath the patella. In the distal third of the lateral side of the leg, the N. fibularis superficialis perforates the fascia to split into the two terminal cutaneous branches (Nn. cutanei dorsalis medialis and intermedius) which continue on the dorsum of the foot. On the posterior side of the leg, the V. saphena parva emerges from the epifascial veins of the lateral margin of the foot and ascends posteriorly to the lateral malleolus on the dorsal side of the calf, pierces the popliteal fascia, and enters the V. poplitea. Adjacent thereof courses the N. cutaneus surae medialis, a branch of the N. tibialis, which continues distally at the distal third of the leg as N. suralis. It frequently communicates with the N. fibularis directly or via a communicating N. cutaneus surae lateralis. The terminal branch of the N. suralis supplies as N. cutaneus dorsalis lateralis the lateral margin of the foot.
Superficial vessels and nerves of the dorsum of the foot
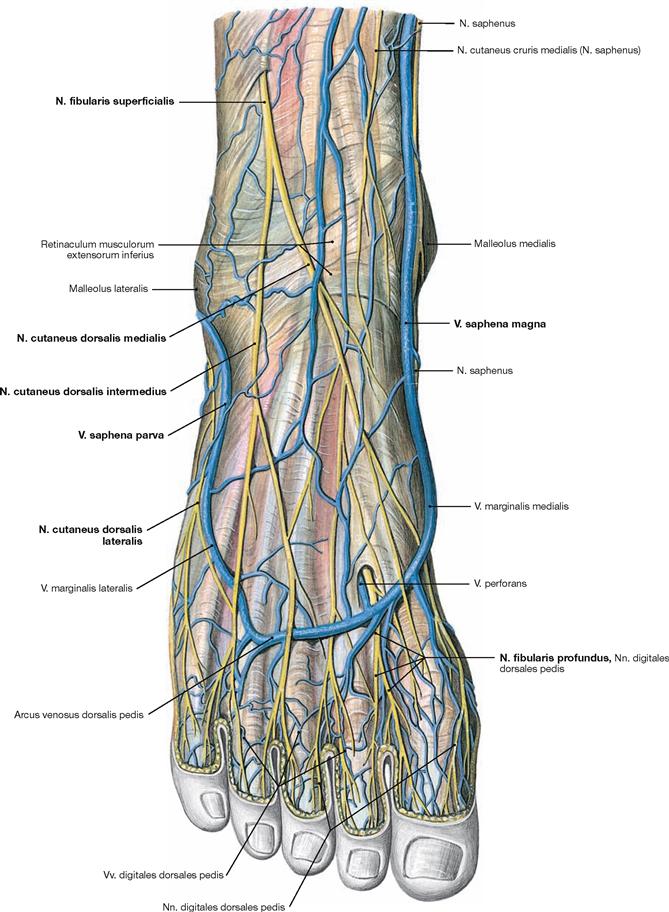
Fig. 4.183 Epifascial veins and nerves of the dorsum of the foot, Dorsum pedis, right side; dorsal view onto the dorsum of the foot.
The V. saphena magna originates at the medial margin of the foot from the epifascial veins of the dorsum of the foot and, thus, is a continuation of the Arcus venosus dorsalis. The smaller V. saphena parva originates from the lateral margin of the foot. At the lateral aspect of the distal leg the N. fibularis superficialis perforates the fascia and splits into the Nn. cutanei dorsales medialis and intermedius for the sensory innervation of the dorsum of the foot and the toes. The lateral margin of the foot is innervated by the N. cutaneus dorsalis lateralis of the N. suralis. Only the first interphalangeal space receives sensory innervation from the terminal branches of the N. fibularis profundus.
Vessels and nerves of the thigh
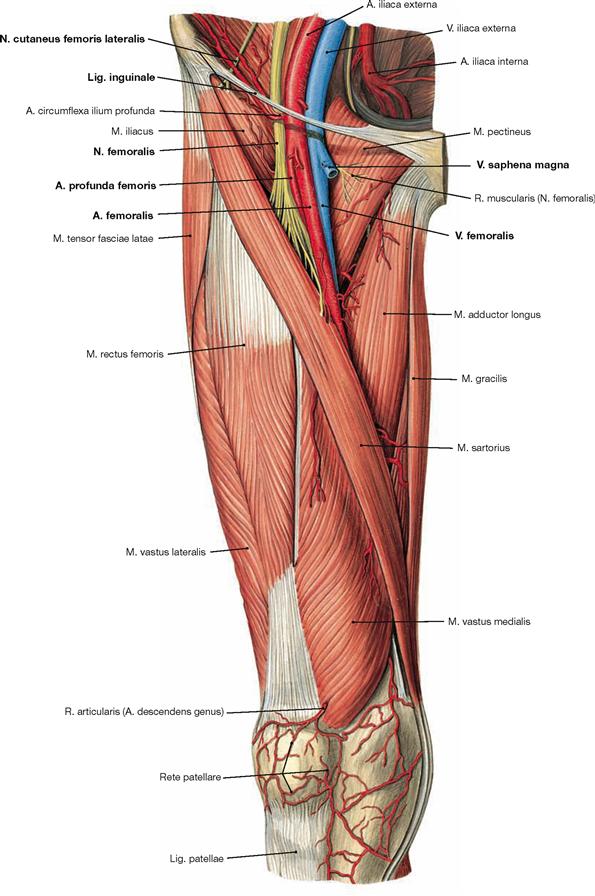
Fig. 4.184 Vessels and nerves of the thigh, Regio femoris anterior, right side; ventral view.
After removal of the fascia, the individual muscles and the subfascial vessels and nerves are displayed in the femoral triangle (Trigonum femorale). The boundaries of the traingle are formed by the inguinal ligament (Lig. inguinale; proximal), the M. gracilis (medial), and the M. sartorius (lateral).
Beneath the inguinal ligament, the following structures enter the triangle from medial to lateral: V. femoralis, A. femoralis, and N. femoralis. The V. saphena magna enters the V. femoralis. The A. femoralis releases smaller branches to the inguinal region 3–6 cm below the inguinal ligament and as a larger branch the A. profunda femoris. In the Fossa iliopectinea, the N. femoralis creates a fan-shaped branching and divides into the N. saphenus, several Rr. musculares for motor innervation of the ventral muscles of the thigh and the M. pectineus as well as the Rr. cutanei anteriores providing sensory innervation to the skin of the ventral thigh. The large cutaneous N. saphenus continues the course of the N. femoralis beneath the M. sartorius. Medial to the Spina iliaca anterior superior, the N. cutaneus femoris lateralis enters the Lacuna musculorum beneath the inguinal ligament.
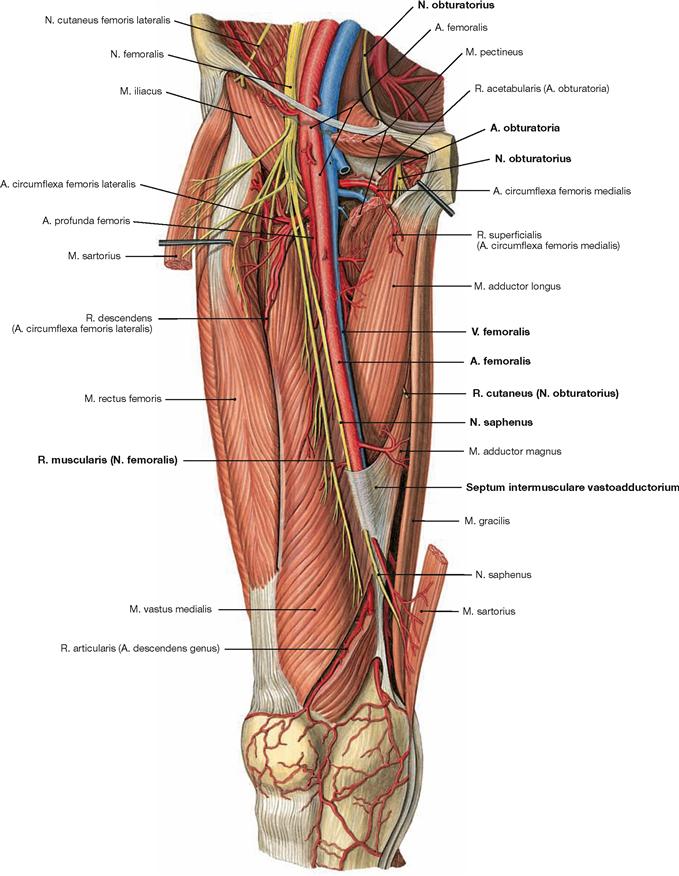
Fig. 4.185 Vessels and nerves of the thigh, Regio femoris anterior, right side; ventral view; after partial removal of the M. sartorius and splitting of the M. pectineus.
The A. and V. femoralis and the N. saphenus are exposed up to their entrance into the adductor canal (Canalis adductorius). The entrance of the adductor canal is formed by the Mm. vastus medialis and adductor longus and the Septum intermusculare vastoadductorium which spans between these muscles and the M. adductor magnus. Owing to the splitting of the M. pectineus, the outlet of the Canalis obturatorius is visible. Here, the N. obturatorius and the A. and V. obturatoria exit the pelvis.
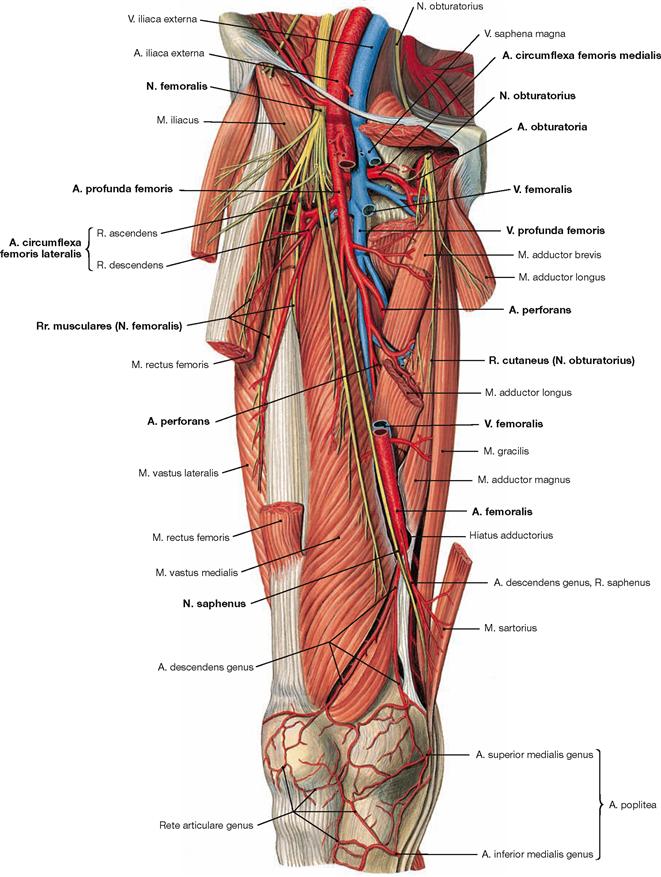
Fig. 4.186 Vessels and nerves of the thigh, Regio femoris anterior, right side; ventral view; after partial removal of the M. sartorius and M. rectus femoris, and splitting of the M. pectineus and M. adductor longus. The adductor canal is opened.
The A. profunda femoris with its branches is displayed. This artery branches off the A. femoralis 3–6 cm below the inguinal ligament and serves as main artery for the thigh and the femoral head (→ pp. 271 and 338). The Aa. circumflexae femoris medialis and lateralis branch off the A. profunda femoris or occasionally derive directly from the A. femoralis. The A. circumflexa femoris medialis has a deep branch to supply the femoral neck and head as well as the adductor muscles and the proximal parts of the hamstring muscles. There are anastomoses with the A. obturatoria which contribute to the supply of the acetabular fossa and the adductors. The ascending branch (R. ascendens) of the A. circumflexa femoris lateralis supplies the lateral muscles of the hip, the descending branch (R. descendens). descendens supplies the anterior ventral muscles of the thigh. The main trunk of the A. profunda femoris descends further and provides three Aa. perforantes to supply the deep adductor muscles and the hamstring muscles at the dorsal aspect of the thigh.
Vessels and nerves of the gluteal region and thigh
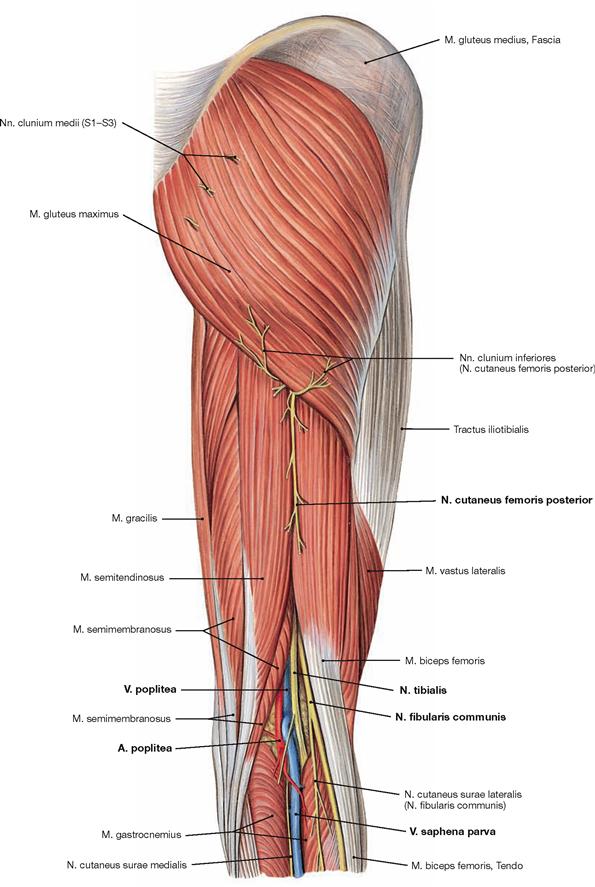
Fig. 4.187 Vessels and nerves of the gluteal region, Regio glutealis, the thigh, Regio femoris posterior, and the popliteal fossa, Fossa poplitea, right side; dorsal view; after removal of the Fascia lata.
The N. cutaneus femoris posterior provides sensory innervation to the posterior aspect of the thigh. It enters the groove between the M. biceps femoris and the M. semitendinosus at the inferior margin of the M. gluteus maximus and penetrates the fascia at the midlength of the thigh. This needs to be considered for the dissection. At the distal thigh, both muscles deviate from each other and define the borders of the popliteal fossa (Fossa poplitea). As a continuation of the A. and V. femoralis, the A. and V. poplitea enter the popliteal fossa after exiting the adductor canal. Here, they are joined by the terminal branches of the N. ischiadicus (N. tibialis and N. fibularis communis). In the popliteal fossa, the N. fibularis communis courses most laterally and superficially. Located medially and deep within the popliteal fossa are the N. tibialis, V. poplitea, and A. poplitea (NVA; from superficial to deep). The V. saphena parva ascends in the middle of the calf and drains into the V. poplitea in the popliteal fossa.
Vessels and nerves of the gluteal region and thigh
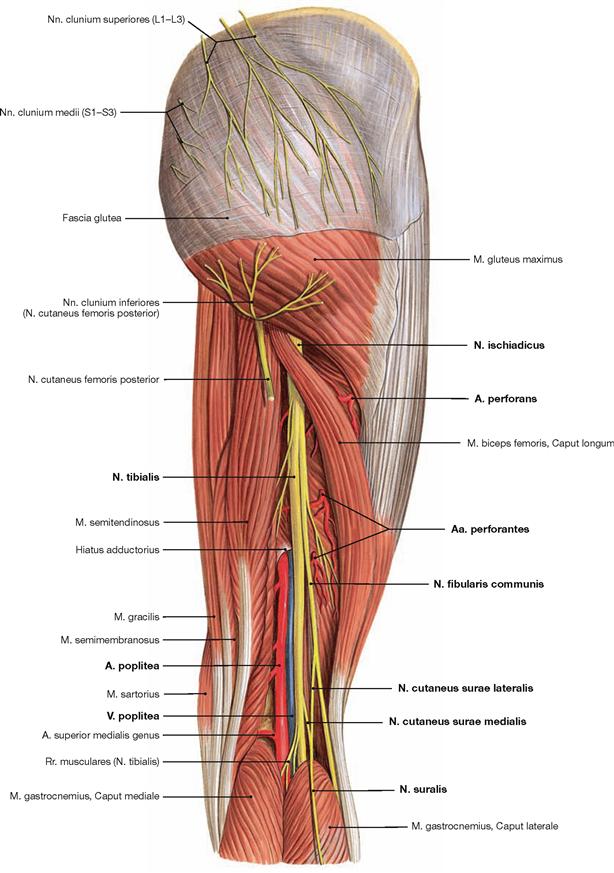
Fig. 4.188 Vessels and nerves of the gluteal region, Regio glutealis, the thigh, Regio femoris posterior, and the popliteal fossa, Fossa poplitea, right side; dorsal view; after removal of the Fascia lata and lateral deflection of the Caput longum of the M. biceps femoris.
The N. ischiadicus descends under the guidance of the M. biceps femoris. At the level of the distal third or higher (as shown here), the N. ischiadicus divides into its terminal branches. The N. tibialis continues its original course and the N. fibularis communis turns laterally to wind around the fibular head and enter the fibularis compartment of the leg below the popliteal fossa. In the region of the popliteal fossa, the N. cutaneus surae medialis branches off the N. tibialis, and the N. cutaneus surae lateralis branches off the N. fibularis communis for the sensory innervation of the calf. The N. cutaneus surae medialis combines with a branch of the N. cutaneus surae lateralis to form the N. suralis. At the thigh, the Aa. perforantes of the A. profunda femoris penetrate the M. adductor magnus lateral to the N. ischiadicus to supply the hamstring muscles.
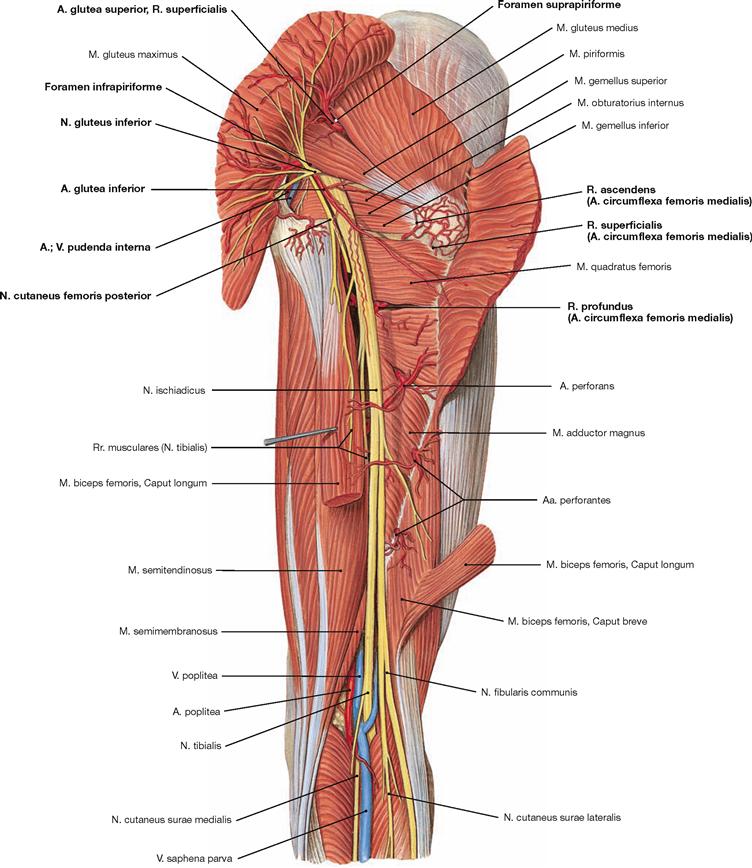
Fig. 4.189 Vessels and nerves of the gluteal region, Regio glutealis, the thigh, Regio femoris posterior, and the popliteal fossa, Fossa poplitea, right side; dorsal view; after dissection of the M. gluteus maximus and Caput longum of the M. biceps femoris.
The N. ischiadicus exits the Foramen infrapiriforme together with the N. cutaneus femoris posterior and the N. gluteus inferior as well as with the A. and V. glutea inferior. The N. pudendus and the A. and V. pudenda interna also exit here, but immediately wind round the Lig. sacrospinale to enter the Fossa ischioanalis beneath the Lig. sacrotuberale through the Foramen ischiadicum minus. The N. gluteus inferior supplies motor fibres to the M. gluteus maximus. The N. gluteus superior together with the A. and V. glutea superior exits the pelvis through the Foramen suprapiriforme but remains at the deep level beneath the M. gluteus medius which it innervates.
Vessels and nerves of the gluteal region
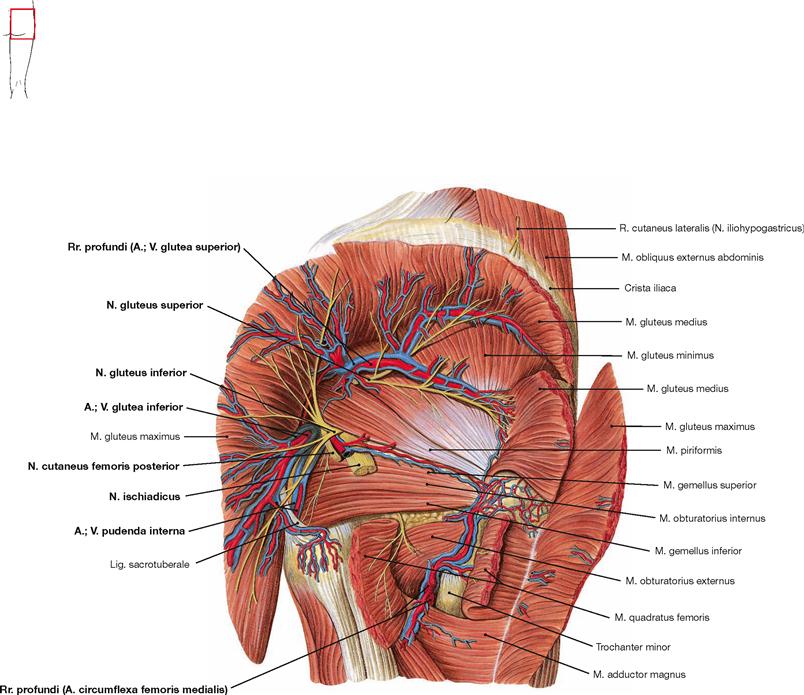
Fig. 4.190 Vessels and nerves of the gluteal region, Regio glutealis, and the thigh, Regio femoris posterior, right side; dorsal view; the Mm. glutei maximus and medius were cut and partially reflected, the N. ischiadicus was removed after its exit from the Foramen infrapiriforme.
After cutting and reflecting the M. gluteus medius, the N. gluteus superior is exposed which exits together with the A. and V. glutea superior through the Foramen suprapiriforme and then courses laterally between the M. gluteus medius and the deeper M. gluteus minimus to the M. tensor fasciae latae. It supplies motor fibres to all these muscles. Several branches of the A. circumflexa femoris medialis appear between the pelvitrochanteric hip muscles and anastomose with the gluteal arteries.
Vessels and nerves of the popliteal fossa
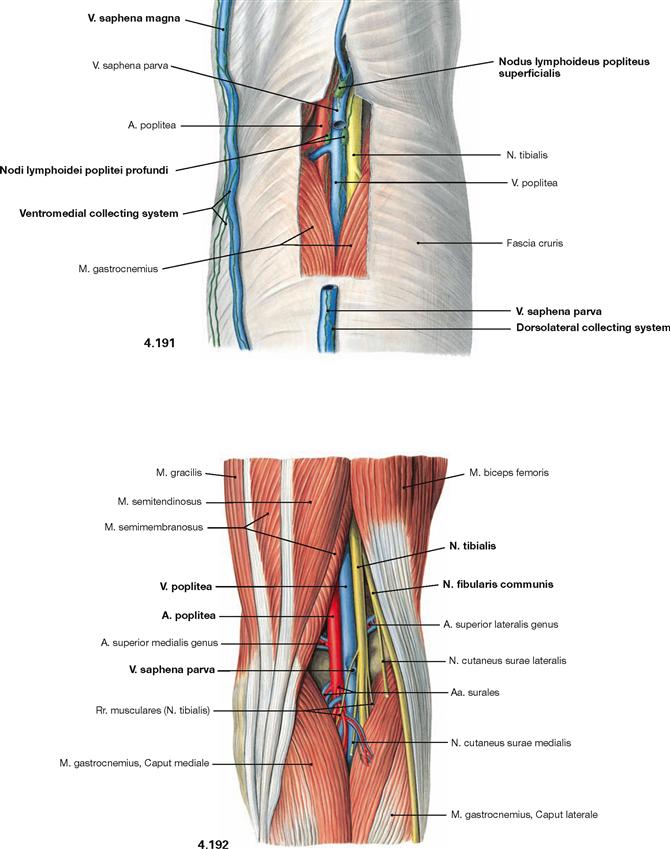
Fig. 4.191 and Fig. 4.192 Vessels and nerves of the popliteal fossa, Fossa poplitea, right side; dorsal view; after partial (→ Fig. 4.191) and complete (→ Fig. 4.192) removal of the fascia.
In the popliteal fossa the N. fibularis communis is located most laterally and superficially, followed medially and deeply by the N. tibialis, V. poplitea, and A. poplitea (NVA). The V. saphena parva ascends in the midline of the leg and drains into the V. poplitea in the popliteal fossa. The dorsolateral lymph vessel system courses along the V. saphena parva, whereas the ventromedial lymph vessel system accompanies the V. saphena magna. The first regional lymph nodes for the dorsolateral collecting system are the Nodi lymphoidei poplitei superficiales and profundi (→ p. 343).
Arteries of the popliteal fossa
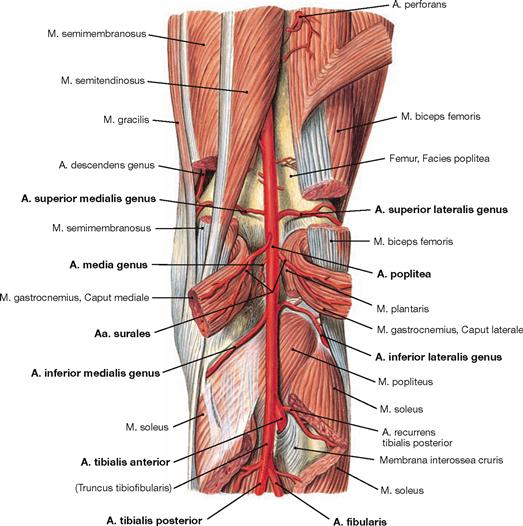
Fig. 4.193 Arteries of the popliteal fossa, Fossa poplitea, right side; dorsal view; after partial resection of the covering muscles.
The A. poplitea supplies the knee joint and forms arterial networks with its branches above (Aa. superiores medialis and lateralis genus) and below (Aa. inferiores medialis and lateralis genus) the articular cavity. These arterial networks contribute to the Rete articulare genus on the ventral side of the knee. At the level of the joint, the A. media genus branches off to supply the knee joint. The Aa. surales supply the muscles of the calf. Below the popliteal fossa, the A. poplitea descends between the two heads of the M. gastrocnemius and divides into the two terminal branches just beneath the tendinous arch of its M. soleus. The A. tibialis posterior continues its course, and the A. tibialis anterior traverses the Membrana interossea cruris to enter the ventral extensor compartment.
Vessels and nerves of the leg
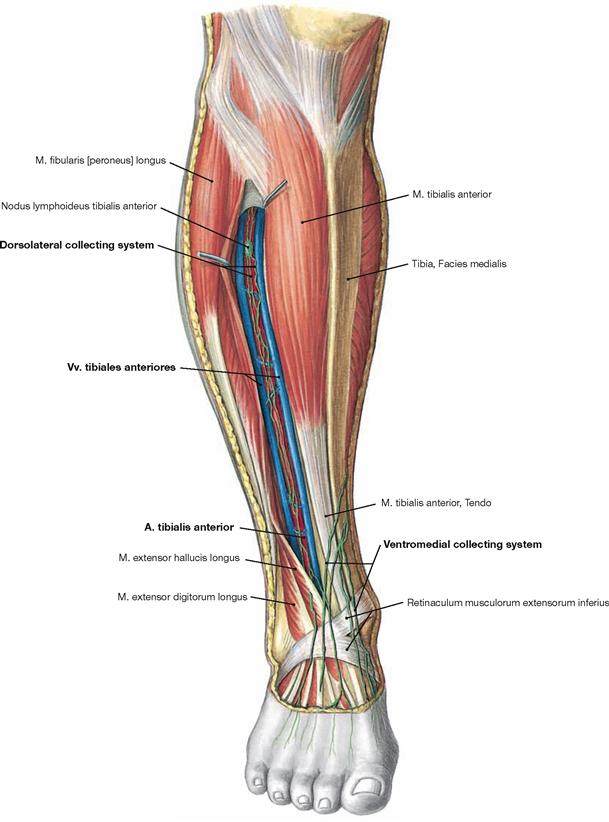
Fig. 4.194 Vessels and nerves of the leg, Regio cruris anterior, right side; ventral view; after spreading apart the extensor muscles.
The superficial lymph vessels course as ventromedial collecting system from the medial margin of the foot alongside the V. saphena magna and as dorsolateral collecting system alongside the V. saphena parva. The deep lymph vessels accompany the arteries in the three muscular compartments as shown here for the extensor compartment.
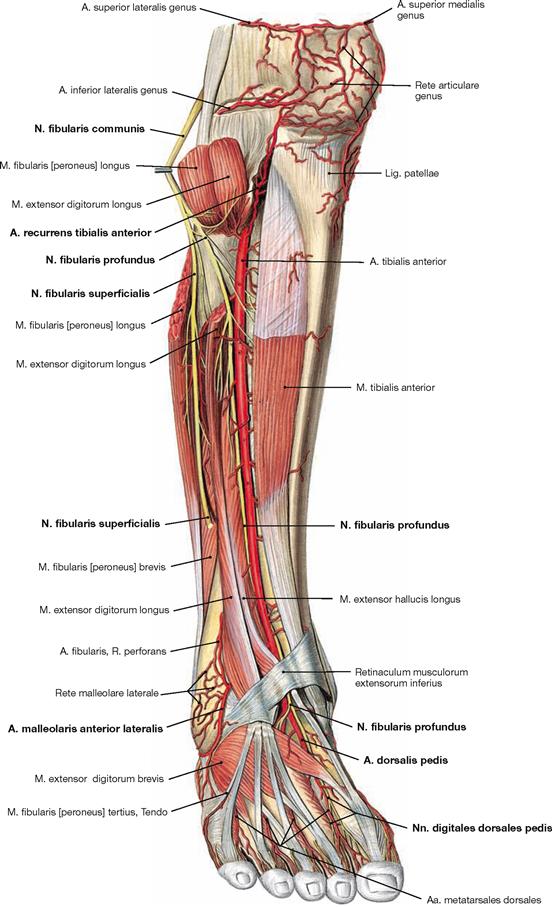
Fig. 4.195 Vessels and nerves of the leg, Regio cruris anterior, right side; ventral view; after removal of the Fascia cruris and dissecting the Mm. extensor digitorum longus and fibularis longus.
The A. tibialis anterior descends in the extensor compartment between the M. extensor digitorum longus and M. tibialis anterior and continues as A. dorsalis pedis on the dorsum of the foot. After the A. recurrens tibialis posterior branches off at the posterior side of the leg, the A. recurrens tibialis anterior appears as the next branch after traversing the Membrana interossea cruris. At the malleoli the Aa. malleolares anteriores medialis and lateralis contribute to the arterial network around the ankle that may provide a sufficient collateral circulation in case of an occlusion of one of the arteries of the leg.
The N. fibularis communis winds laterally around the head of the fibula, enters the fibularis compartment, and then divides into its two terminal branches. The N. fibularis superficialis descends in the fibularis compartment, provides motor fibres to both fibularis muscles and pierces the fascia at the distal third of the leg. The N. fibularis profundus passes over to the extensor compartment and descends adjacent to the A. tibialis anterior. It conveys motor innervation to all extensors of the leg and the dorsum of the foot. Its terminal branches convey sensory fibres to the first interdigital space.
Vessels and nerves of the popliteal fossa and leg
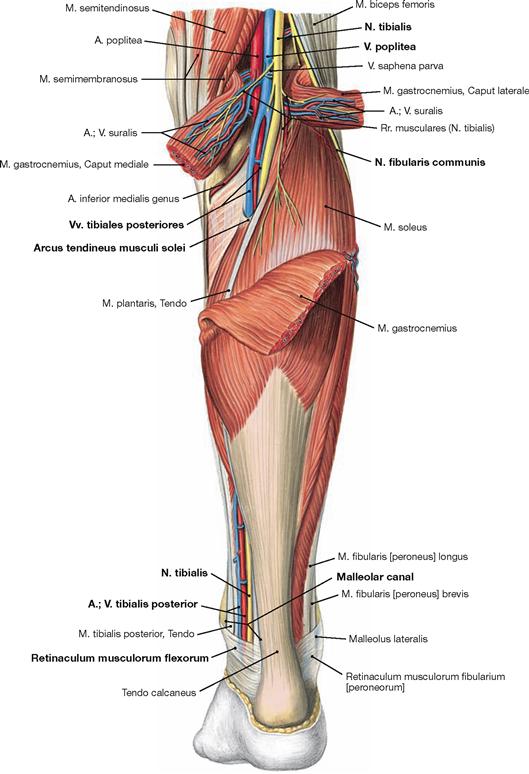
Fig. 4.196 Vessels and nerves of the popliteal fossa, Fossa poplitea, and the leg, Regio cruris posterior, right side; dorsal view; after removal of the Fascia cruris and dissecting the M. gastrocnemius.
Joined by two concomitant veins and the N. tibialis, the A. tibialis posterior courses beneath the tendinous arch of the M. soleus (Arcus tendineus musculi solei) and descends between the superficial and deep flexor muscles of the leg to the medial malleolus. There, it traverses the malleolar canal beneath the Retinaculum musculorum flexorum to reach the sole of the foot.
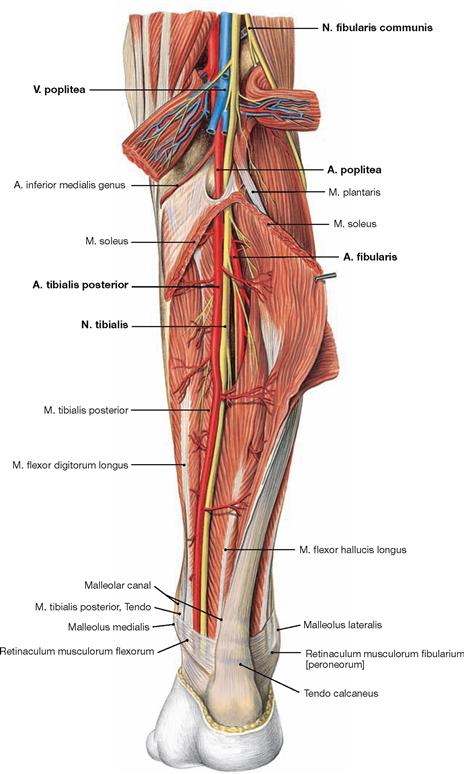
Fig. 4.197 Vessels and nerves of the popliteal fossa, Fossa poplitea, and the leg, Regio cruris posterior, right side; dorsal view; after dissecting the Mm. gastrocnemius and soleus.
Shortly after passing through the tendinous arch of the M. soleus, the A. tibialis posterior gives rise to its most important branch, the A. fibularis, which descends to the lateral malleolus.
Vessels and nerves of the leg
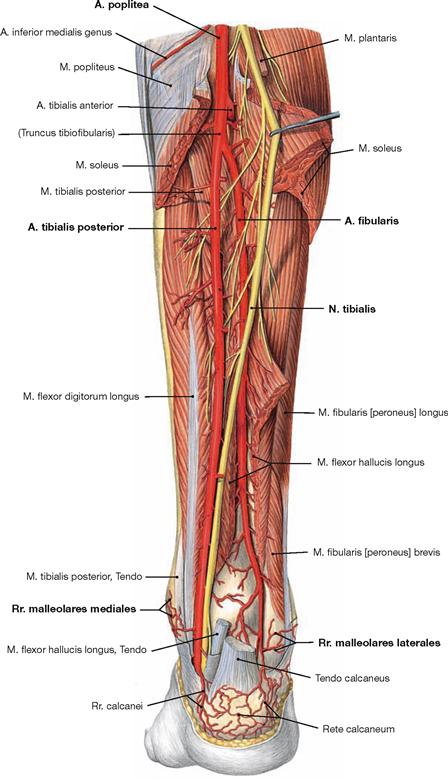
Fig. 4.198 Vessels and nerves of the leg, Regio cruris posterior, right side; dorsal view; after removal of the Fascia cruris and dissecting the Mm. gastrocnemius, soleus, and flexor hallucis longus.
The A. tibialis posterior descends together with the N. tibialis between the superficial and deep flexor muscles of the leg to the medial malleolus and continues through the malleolar canal beneath the Retinaculum musculorum flexorum to the plantar side of the foot. Rr. malleolares mediales to the medial malleolus derive from this vessel.
The A. fibularis pierces the M. flexor hallucis longus and descends in the deepest layer directly on the Membrana interossea cruris to the lateral malleolus. Together with the branches of the Aa. tibiales anterior and posterior, its Rr. malleolares laterales complete the arterial network around the ankle which provides sufficient collaterals to compensate for an occluded vessel.
Vessels and nerves of the dorsum of the foot
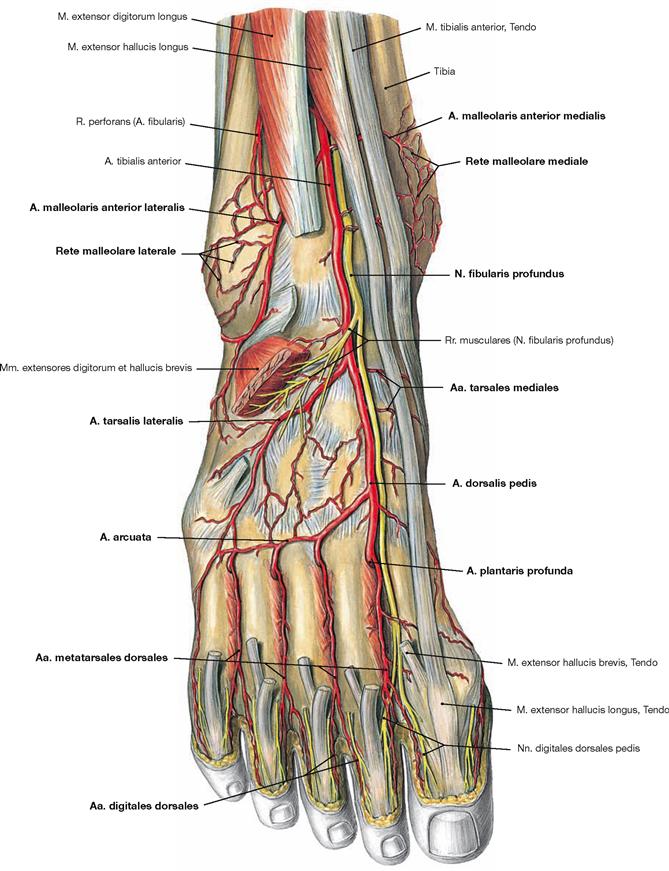
Fig. 4.199 Vessels and nerves of the dorsum of the foot, Dorsum pedis, right side; dorsal view onto the dorsum of the foot after removal of the tendons of the M. extensor digitorum longus and the short extensors of the toes.
The A. tibialis anterior continues on the dorsum of the foot as A. dorsalis pedis. After the innervation of the extensors of the leg and the dorsum of the foot, the concomitant N. fibularis profundus divides into terminal sensory branches which supply the first interdigital space. At the level of the malleoli, the A. tibialis anterior provides the Aa. malleolares anteriores medialis and lateralis for the arterial networks around the malleoli (Rete malleolare mediale and Rete malleolare laterale). The A. dorsalis pedis provides several smaller Aa. tarsales mediales and one A. tarsalis lateralis to the Tarsus and then continues as A. arcuata. The latter arches to the lateral margin of the foot and gives rise to the Aa. metatarsales dorsales which continue as Aa. digitales dorsales to supply the toes. The A. plantaris profunda participates in the perfusion of the sole of the foot by supplying the Arcus plantaris profundus.
Arteries of the sole of the foot
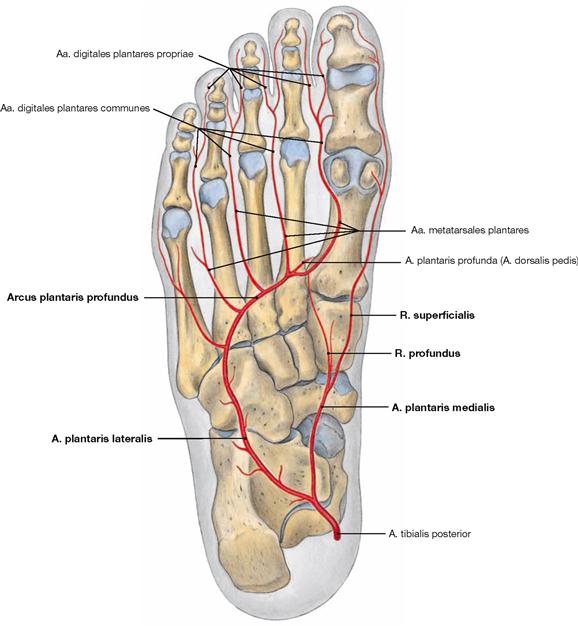
Fig. 4.200 Arteries of the sole of the foot, Planta pedis, right side; plantar view.
The plantar aspect of the foot is perfused by terminal branches of the A. tibialis posterior. The A. plantaris medialis provides a R. superficialis to the medial margin of the foot and a R. profundus which connects to the Arcus plantaris profundus. This arterial arch is a direct continuation of the A. plantaris lateralis.
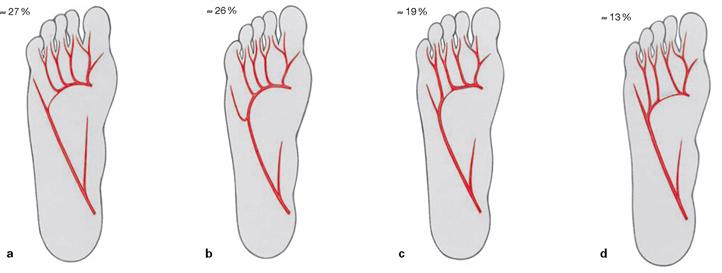
Figs. 4.201a to d Variations of the arterial supply of the toes, right side; plantar view.
The Arcus plantaris profundus may receive its main input from the A. dorsalis pedis via the A. plantaris profunda (a) or from the A. tibialis posterior (b). Alternatively, both arteries may contribute to the arterial supply of the toes (c and d).
Vessels and nerves of the sole of the foot
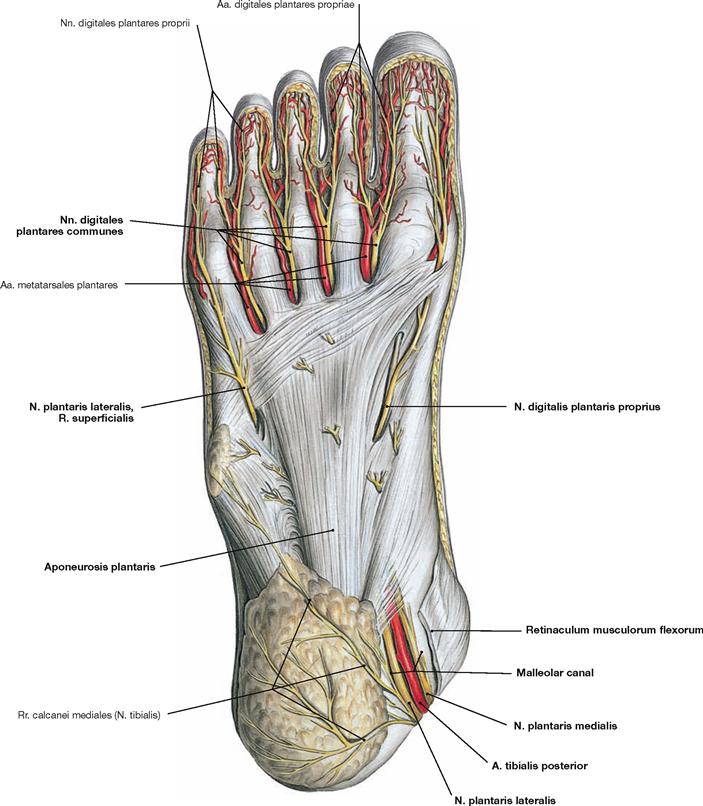
Fig. 4.202 Superficial layer of the arteries and nerves of the sole of the foot, right side; plantar view.
The N. tibialis already divides into its two terminal branches (Nn. plantares medialis and lateralis) at the medial malleolus within the malleolar canal beneath the Retinaculum musculorum flexorum. The terminal branches then divide further into several Nn. digitales plantares. Similar to the N. ulnaris at the hand, the N. plantaris lateralis divides into a R. superficialis and a R. profundus. The N. plantaris medialis supplies an additional N. digitalis plantaris proprius at the medial margin of the foot. The sensory branches surface between the longitudinal fibres of the plantar aponeurosis (Aponeurosis plantaris). The A. tibialis posterior divides only at the plantar aspect of the foot.
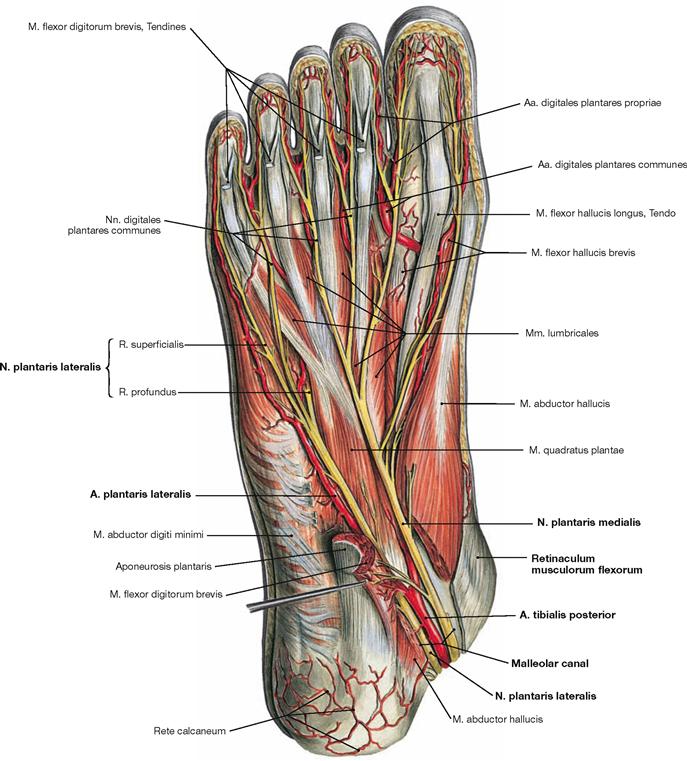
Fig. 4.203 Intermediate layer of the arteries and nerves of the sole of the foot, right side; plantar view.
The M. flexor digitorum brevis and the M. abductor hallucis were cut to expose the neurovascular passageway of the malleolar canal. The Nn. plantares medialis and lateralis are accompanied by the corresponding vessels from the A. tibialis posterior. The blood vessels continue beneath the M. flexor digitorum brevis to reach the intermediate layer of the neurovascular structures to the toes. On their way, the nerves provide motor fibres to the short muscles of the sole of the foot.
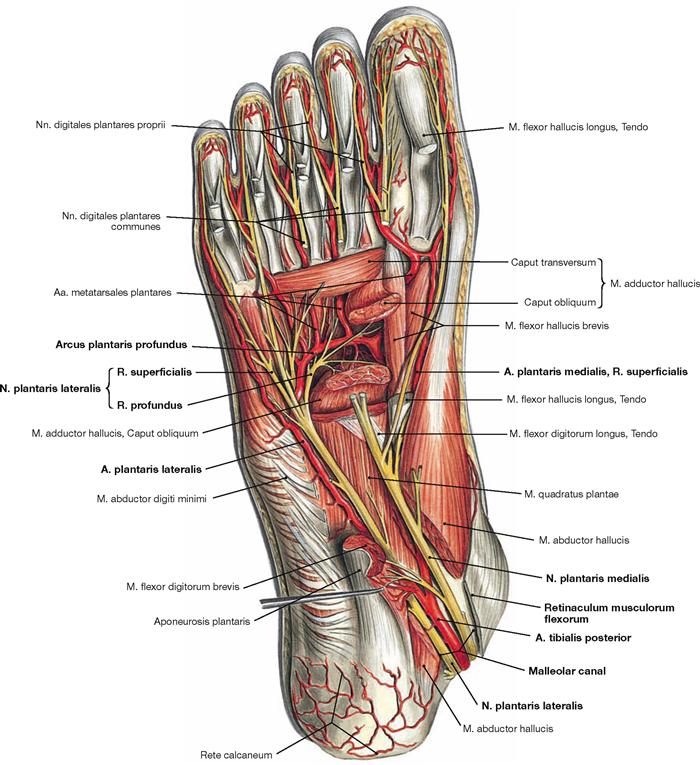
Fig. 4.204 Deep layer of the arteries and nerves of the sole of the foot, right side; plantar view.
The M. flexor digitorum brevis and the M. abductor hallucis were split to expose the neurovascular passageway of the malleolar canal. In addition, the Caput obliquum of the M. adductor hallucis was cut to demonstrate the deep plantar arch (Arcus plantaris profundus) and the course of the R. profundus of the N. plantaris lateralis. The Arcus plantaris profundus continues the A. plantaris lateralis und receives blood from the R. profundus of the A. plantaris medialis and from the A. plantaris profunda which derives from the A. dorsalis pedis. Together with the R. profundus from the N. plantaris lateralis it arches over the Mm. interossei of the sole of the foot in the deep layer of the neurovascular structures.
Foot, sagittal sections
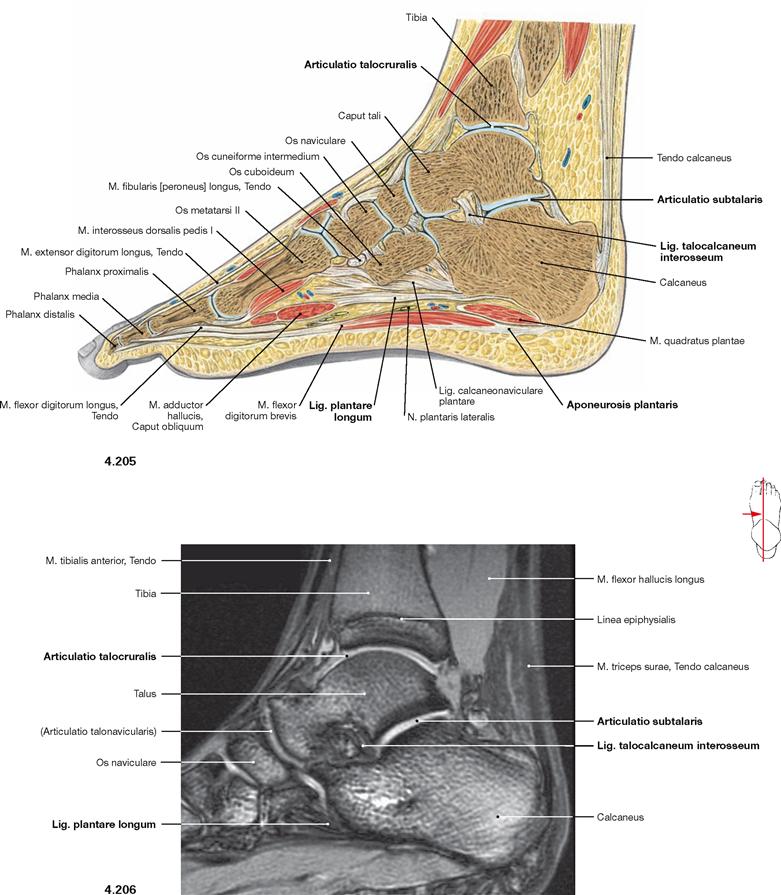
Fig. 4.205 and Fig. 4.206 Foot, Pes, right side; sagittal section through the second phalanx (→ Fig. 4.205) and corresponding magnetic resonance imaging (MRI) sagittal section (→ Fig. 4.206); medial view.
The section visualises the articular cavity of the ankle joint (Articulatio talocruralis) and the posterior chamber of the talocalcaneonavicular joint (Articulatio subtalaris). The longitudinal arch is stabilised by three overlying ligamentous systems (Aponeurosis plantaris, Lig. plantare longum, Lig. calcaneonaviculare plantare) (→ Fig. 4.95).
Hip joint, oblique section
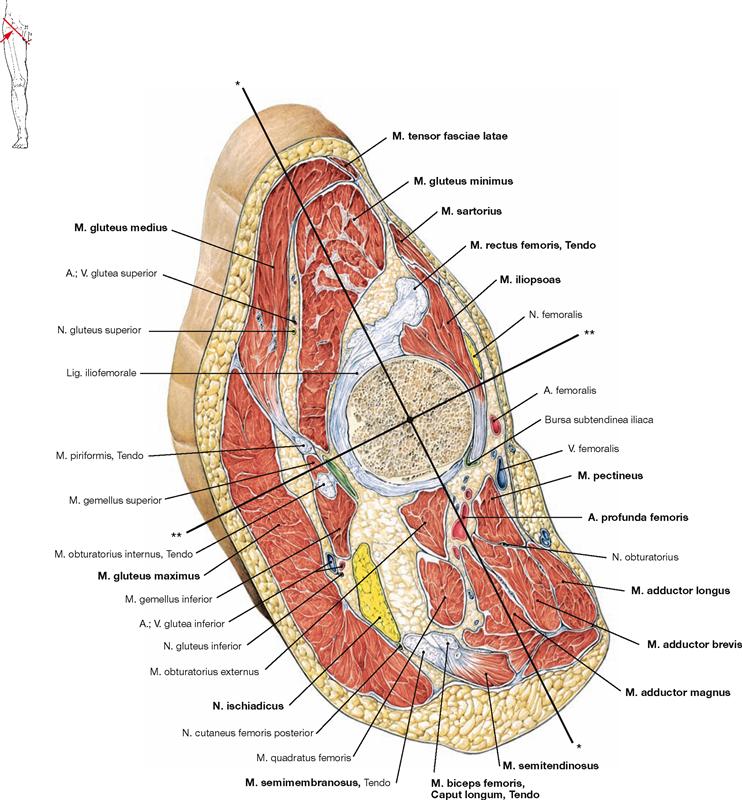
Fig. 4.207 Thigh, Femur, oblique section through the hip joint, right side; distal view illustrating the axes of movement of the hip joint.
The oblique section through the thigh at the level of the femoral head shows the position of the diverse groups of muscles relative to the articular head and the axes of movement. The M. gluteus maximus is located dorsal to the hip joint, whereas the smaller gluteal muscles (Mm. glutei medius and minimus) in part course ventral to the longitudinal and transverse axes of the hip joint. This position explains why the M. gluteus maximus acts as external rotator and extensor of the hip, and the small gluteal muscles function as strongest medial rotators and also as flexors of the hip. The M. iliopsoas is located anterior to the transverse axis and is the most important flexor of the hip joint. It is supported for this function by the anterior group of femoral muscles (M. sartorius, M. rectus femoris), the M. tensor fasciae latae, and the superficial adductor muscles (Mm. adductores longus and brevis, M. pectineus, main part of the M. adductor magnus). However, the dorsal part of the M. adductor magnus is positioned posterior to the transverse axis and functions as extensor of the hip joint together with the hamstring muscles of which it is a part of given its function and innervation. Cross-sections through the extremities are well suited to comprehend the course of the neurovascular structures in the respective compartments at several levels. After exiting the small pelvis, the N. ischiadicus initially courses beneath the M. gluteus maximus. On the ventral side, the A. profunda femoris is covered by the M. pectineus.
* transverse axis of movement in the hip joint
** sagittal axis of movement in the hip joint
Sections
Thigh, transverse section
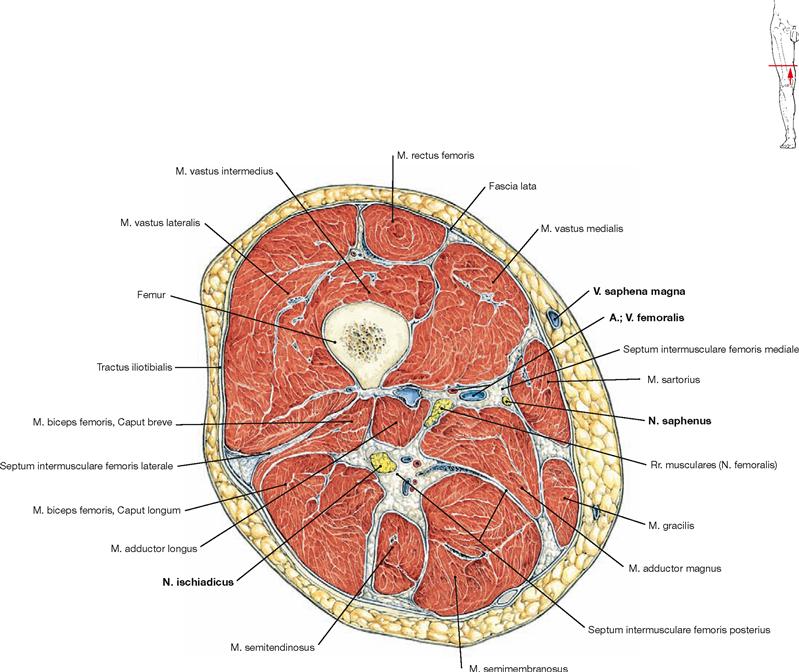
Fig. 4.208 Thigh, Femur, right side; transverse section at the mid-tigh level; distal view.
This cross-section shows the three muscle groups of the thigh. The ventral group comprises the M. quadriceps femoris and the M. sartorius. Medially located are the adductor muscles and dorsally the hamstring muscles.
The V. saphena magna is found in the epifascial subcutaneous adipose tissue on the medial aspect of the thigh. The A. and V. femoralis jointly course with the N. saphenus through the adductor canal (Canalis adductorius) of the M. quadriceps. The adductor canal is demarcated dorsally by the Mm. adductores longus and magnus, medially by the M. vastus medialis, and ventrally by the M. sartorius. The N. ischiadicus is positioned dorsally beneath the M. biceps femoris.
Knee, transverse section
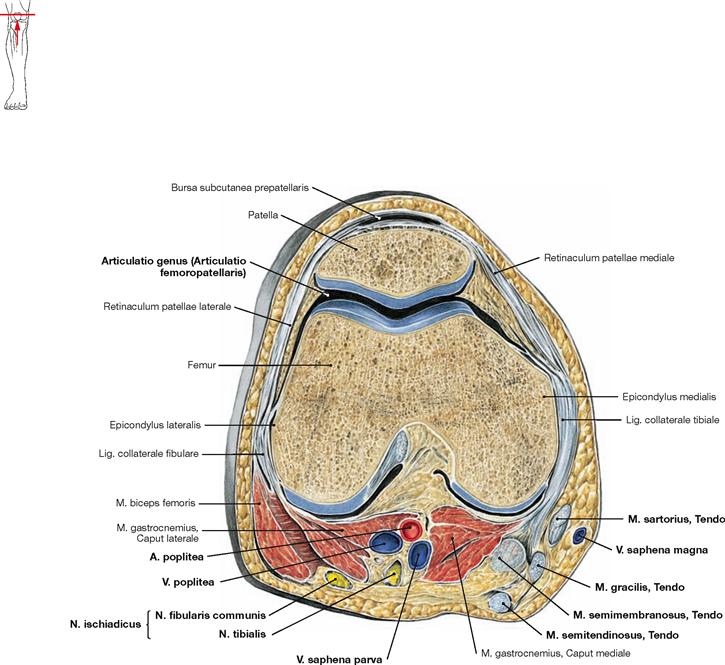
Fig. 4.209 Knee joint, Articulatio genus, right side; transverse section; distal view.
The transverse section through the knee joint shows the articular surfaces of the Articulatio femoropatellaris. On the posterior side, the M. biceps femoris is positioned laterally. Therefore this muscle is the most important lateral rotator. On the medial side, several muscles contribute to medial rotation. The tendons of the Mm. sartorius, gracilis, and semitendinosus are located superficially. They insert further distally with a common aponeurosis at the medial aspect of the Tibia, commonly referred to as “Pes anserinus superficialis”. Beneath it, the insertion of the M. semimembranosus tendon is referred to as “Pes anserinus profundus”.
The V. saphena magna is found in the epifascial subcutaneous adipose tissue on the medial side of the knee. Dorsally, the terminal branches of the N. ischiadicus (N. tibialis and N. fibularis communis) are the most superficial structures, followed further beneath by the V. poplitea with the confluence of the V. saphena parva, and deepest within the popliteal fossa the A. poplitea (NVA).
Leg, transverse section
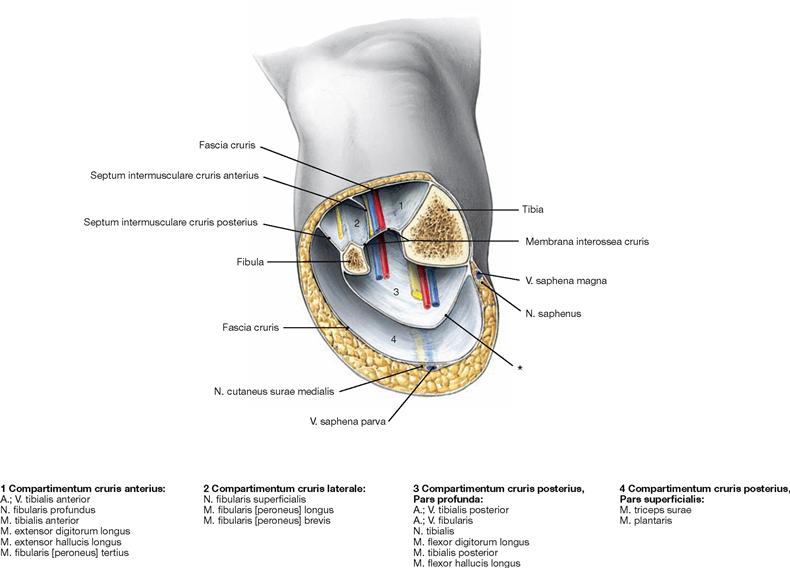
Fig. 4.210 Leg, Crus, right side; transverse section at the mid-leg level with illustration of the osteofibrous compartments; distal view.
The Fascia cruris is attached to the bones of the leg by dense connective tissue septa. They separate osteofibrous compartments in which the neurovascular structures are embedded between the respective muscle groups (→ Fig. 4.211). The Septum intermusculare anterius partitions the extensor compartment anteriorly from the lateral fibularis compartment which in turn is separated from the superficial flexors by the Septum intermusculare posterius. The superficial flexors are isolated from the deep flexors by a deep layer of the Fascia cruris.
The deep flexors are directly adjacent to the Membrana interossea cruris. The anterior (extensor) compartment harbours the N. fibularis profundus, the A. tibialis and the Vv. tibiales anteriores. The N. fibularis superficialis is located in the lateral (fibularis) compartment. In the deep posterior (flexor) compartment, the N. tibialis, A. tibialis posterior, Vv. tibialis posteriores, and – covered by the M. flexor hallucis longus – the A. and V. fibularis are embedded in the muscles. The V. saphena magna and V. saphena parva on the dorsal side course in the epifascial layer at the medial aspect of the leg.
* deep part of the Fascia cruris
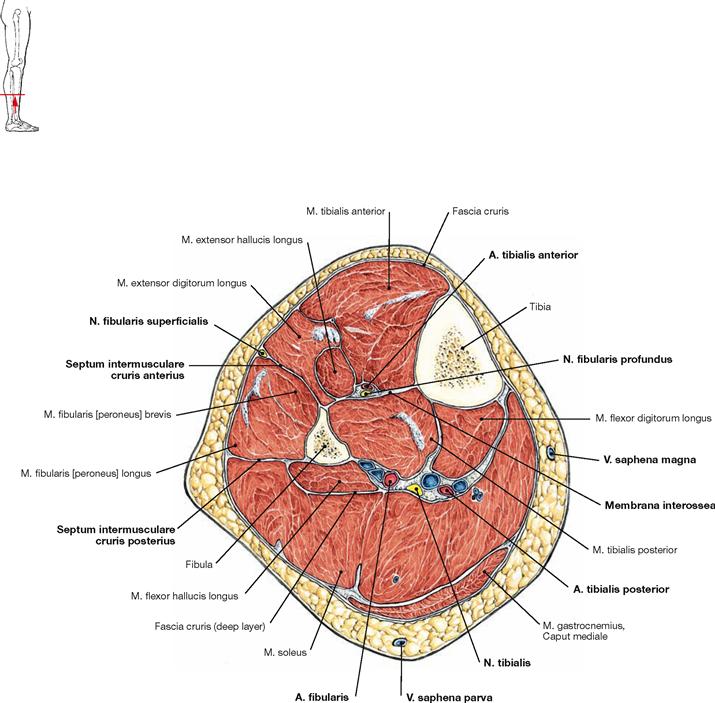
Fig. 4.211 Leg, Crus, right side; transverse section at the mid-leg level, distal view.
Together with the connective tissue septa reaching to the bones of the leg, the Fascia cruris confines the osteofibrous compartments. These compartments contain the respective neurovascular structures embedded between the muscle bellies of the individual muscle groups.
Of highest clinical relevance is the anterior (extensor) compartment which harbours the N. fibularis profundus together with the A. tibialis anterior.